

Revisiting the Plasmodium falciparum druggable genome using predicted structures and data mining
- PMID: 40066064
- PMCID: PMC11892419
- DOI: 10.1038/s44386-025-00006-5
Revisiting the Plasmodium falciparum druggable genome using predicted structures and data mining
Abstract
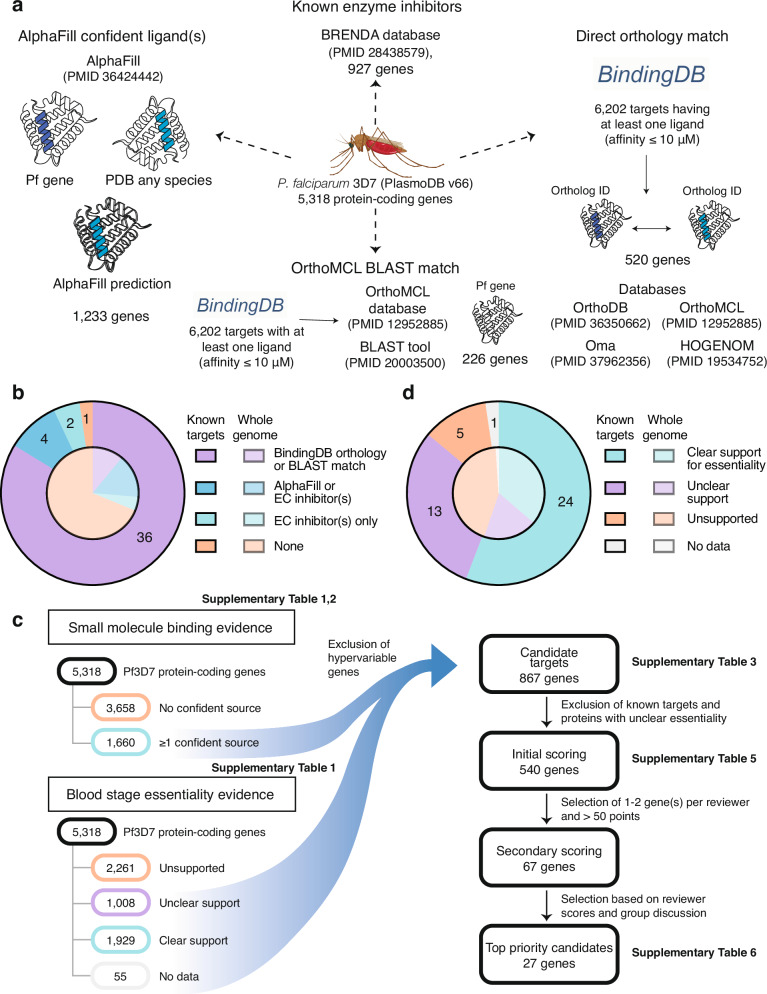
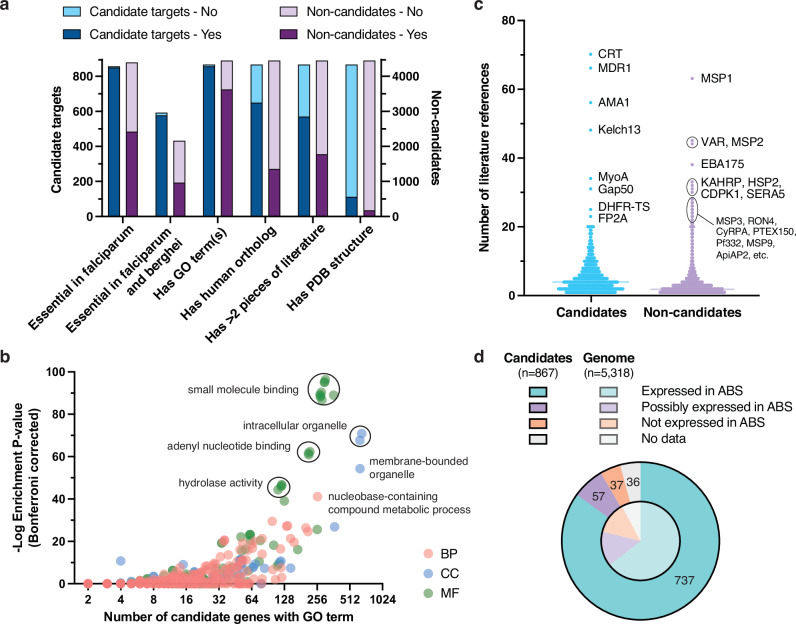
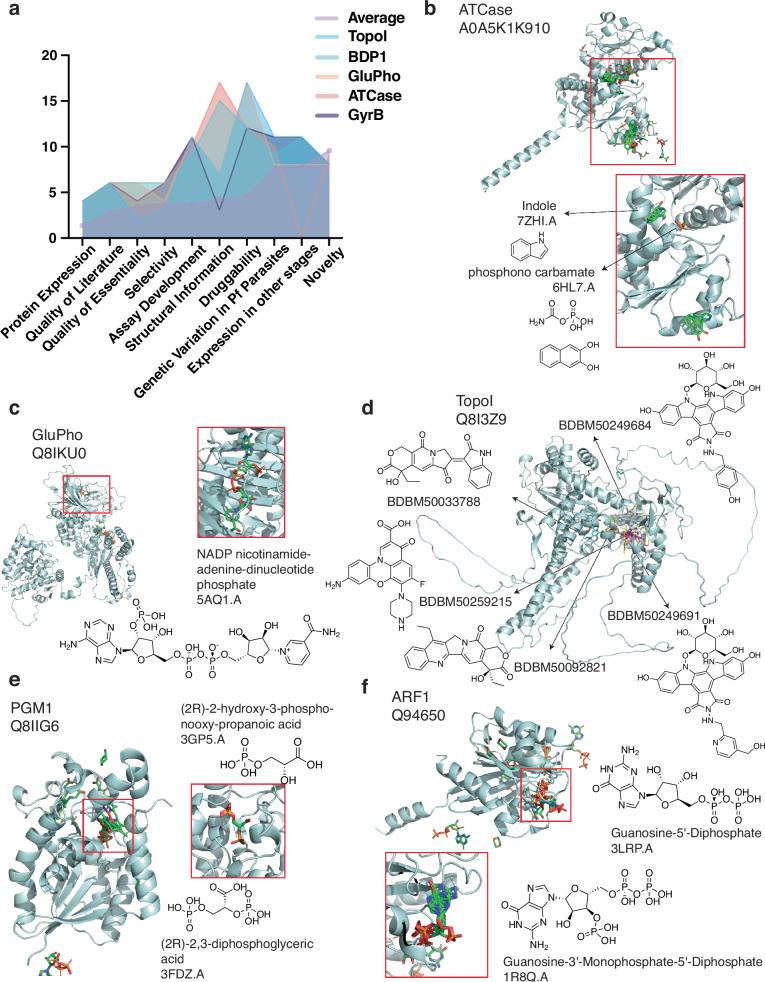
Identification of novel drug targets is a key component of modern drug discovery. While antimalarial targets are often identified through the mechanism of action studies on phenotypically derived inhibitors, this method tends to be time- and resource-consuming. The discoverable target space is also constrained by existing compound libraries and phenotypic assay conditions. Leveraging recent advances in protein structure prediction, we systematically assessed the Plasmodium falciparum genome and identified 867 candidate protein targets with evidence of small-molecule binding and blood-stage essentiality. Of these, 540 proteins showed strong essentiality evidence and lack inhibitors that have progressed to clinical trials. Expert review and rubric-based scoring of this subset based on additional criteria such as selectivity, structural information, and assay developability yielded 27 high-priority antimalarial target candidates. This study also provides a genome-wide data resource for P. falciparum and implements a generalizable framework for systematically evaluating and prioritizing novel pathogenic disease targets.
Keywords: Target identification; Target validation.
© The Author(s) 2025.
Conflict of interest statement
Competing interestsM.K.G. has an equity interest in and is a cofounder and scientific advisor of VeraChem LLC, and is on the SABs of InCerebro Inc, Denovicon Therapeutics, and Beren Therapeutics. E.L.F. and S.P.S.R. are employees of Novartis Pharma AG and may own shares in Novartis Pharma AG. K.D. holds stock in TropIQ Health Sciences. The remaining authors declare no competing interests.
Figures

Update of
-
Revisiting the Plasmodium falciparum druggable genome using predicted structures and data mining.Res Sq [Preprint]. 2024 Nov 26:rs.3.rs-5412515. doi: 10.21203/rs.3.rs-5412515/v1. Res Sq. 2024. Update in: NPJ Drug Discov. 2025;2(1):3. doi: 10.1038/s44386-025-00006-5. PMID: 39649165 Free PMC article. Updated. Preprint.
References
Grants and funding
LinkOut - more resources
Full Text Sources