

Tuft cell IL-17RB restrains IL-25 bioavailability and reveals context-dependent ILC2 hypoproliferation
- PMID: 40074948
- PMCID: PMC11957993
- DOI: 10.1038/s41590-025-02104-y
Tuft cell IL-17RB restrains IL-25 bioavailability and reveals context-dependent ILC2 hypoproliferation
Abstract
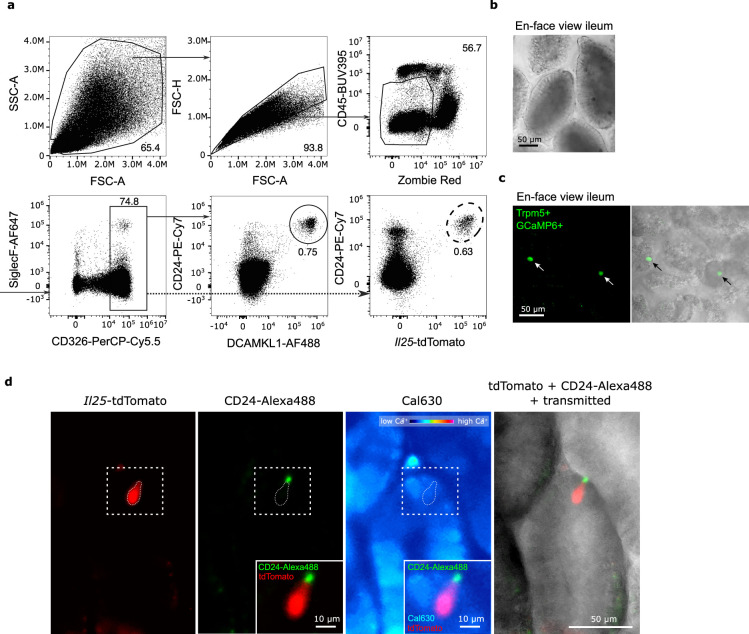
The tuft cell-group 2 innate lymphoid cell (ILC2) circuit orchestrates rapid type 2 responses upon detecting microbially derived succinate and luminal helminths. Our findings delineate key mechanistic steps involving IP3R2 engagement and Ca2+ flux, governing interleukin-25 (IL-25) production by tuft cells triggered by succinate detection. While IL-17RB has a pivotal intrinsic role in ILC2 activation, it exerts a regulatory function in tuft cells. Tuft cells exhibit constitutive Il25 expression, placing them in an anticipatory state that facilitates rapid production of IL-25 protein for ILC2 activation. Tuft cell IL-17RB is crucial for restraining IL-25 bioavailability, preventing excessive tonic ILC2 stimulation due to basal Il25 expression. Supraoptimal ILC2 stimulation by IL-25 resulting from tuft cell Il17rb deficiency or prolonged succinate exposure induces a state of hypoproliferation in ILC2s, also observed in chronic helminth infection. Our study offers critical insights into the regulatory dynamics of IL-25 in this circuit, highlighting the delicate tuning required for responses to diverse luminal states.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures













Update of
-
Tuft cell IL-17RB restrains IL-25 bioavailability and reveals context-dependent ILC2 hypoproliferation.bioRxiv [Preprint]. 2024 Mar 8:2024.03.04.583299. doi: 10.1101/2024.03.04.583299. bioRxiv. 2024. Update in: Nat Immunol. 2025 Apr;26(4):567-581. doi: 10.1038/s41590-025-02104-y. PMID: 38496438 Free PMC article. Updated. Preprint.
References
-
- Gerbe, F. & Jay, P. Intestinal tuft cells: epithelial sentinels linking luminal cues to the immune system. Mucosal Immunol.9, 1353–1359 (2016). - PubMed
MeSH terms
Substances
Grants and funding
- R01 AR070116/AR/NIAMS NIH HHS/United States
- R01 AI095466/AI/NIAID NIH HHS/United States
- SPP1937 - KL 2963/3-1 and KL 2963/2-1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- CRC/TRR 241 Project-ID 375876048/Deutsche Forschungsgemeinschaft (German Research Foundation)
- R01 DK132244/DK/NIDDK NIH HHS/United States
- R01 DK126871/DK/NIDDK NIH HHS/United States
- Sonderforschungsbereich-Transregio TRR 152/Deutsche Forschungsgemeinschaft (German Research Foundation)
- TRR 152/Deutsche Forschungsgemeinschaft (German Research Foundation)
- R01 AI151599/AI/NIAID NIH HHS/United States
- 194216/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- U01 AI095608/AI/NIAID NIH HHS/United States
- R01 AI172027/AI/NIAID NIH HHS/United States
- FOR2599 Project-ID 22359157/Deutsche Forschungsgemeinschaft (German Research Foundation)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
