

This is a preprint.
Topographic organization of bidirectional connections between the cingulate region (infralimbic area and anterior cingulate area, dorsal part) and the interbrain (diencephalon) of the adult male rat
- PMID: 40093037
- PMCID: PMC11908189
- DOI: 10.1101/2024.09.29.615708
Topographic organization of bidirectional connections between the cingulate region (infralimbic area and anterior cingulate area, dorsal part) and the interbrain (diencephalon) of the adult male rat
Abstract
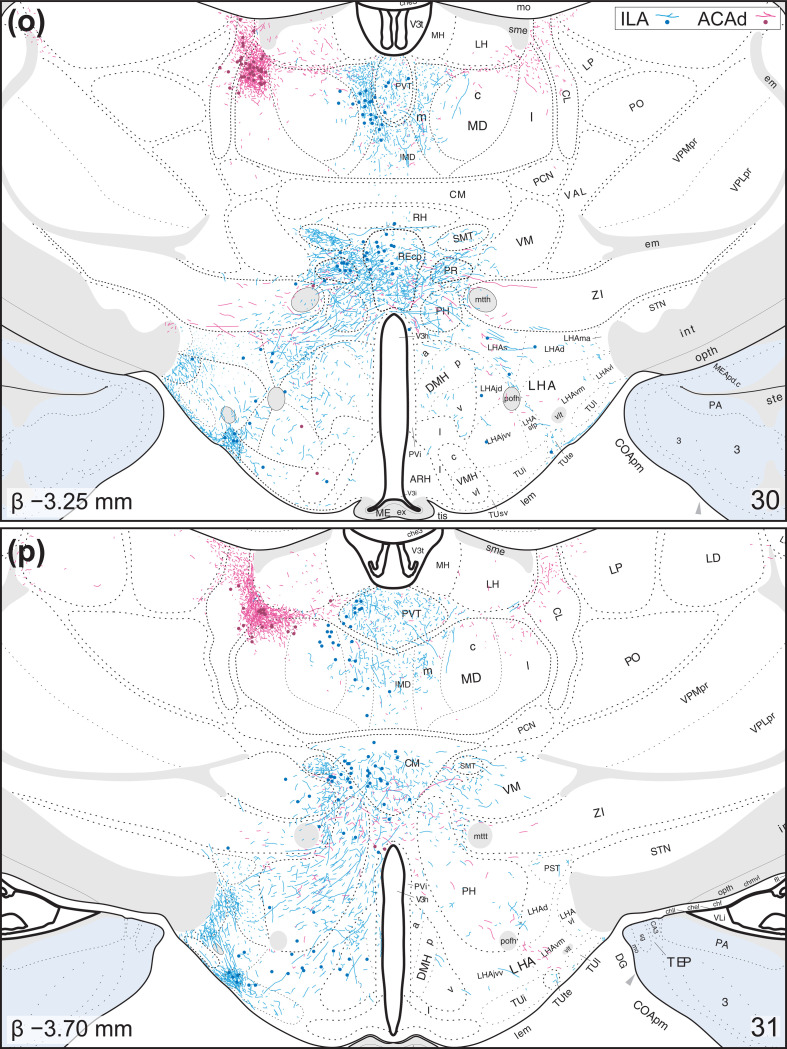
The medial prefrontal cortex [cingulate region (Brodmann, 1909) (CNG)] in the rat is a connectionally and functionally diverse structure. It harbors cerebral nuclei that use long-range connections to promote adaptive changes to ongoing behaviors. The CNG is often described across functional and anatomical gradients, a dorsalventral gradient being the most prominent. Topographic organization is a general feature of the nervous system, and it is becoming clear that such spatial arrangements can reflect connectional, functional, and cellular differences. Portions of the CNG are known to form reciprocal connections with cortical areas and thalamus; however, these connectional features have not been described in detail or mapped to standardized rat brain atlases. Here, we used co-injected anterograde (Phaseolus vulgaris leucoagglutinin) and retrograde (cholera toxin B subunit) tracers throughout the CNG to identify zones of reciprocal connectivity in the diencephalon [or interbrain (Baer, 1837) (IB)]. Tracer distributions were observed using a Nissl-based atlas-mapping approach that facilitates description of topographic organization. This draft report describes CNG connections of the infralimbic area (Rose & Woolsey, 1948) (ILA) and the anterior cingulate area, dorsal part (Krettek & Price, 1977) (ACAd) throughout the IB. We found that corticothalamic connections are predominantly reciprocal, and that ILA and ACAd connections tended to be spatially segregated with minimal overlap. In the hypothalamus (Kuhlenbeck, 1927), we found dense and specific ILA-originating terminals in the following Brain Maps 4.0 atlas territories: dorsal region (Swanson, 2004) (LHAd) and suprafornical region (Swanson, 2004) (LHAs) of the lateral hypothalamic area (Nissl, 1913), parasubthalamic nucleus (Wang & Zhang, 1995) (PSTN), tuberal nucleus, terete part (Petrovich et al., 2001) (TUte), and an ill-defined dorsal cap of the medial mammillary nucleus (Gudden, 1881) (MM). We discuss these findings in the context of feeding behaviors.
Keywords: anterograde; brain atlas; feeding; lateral hypothalamic area; mapping; prefrontal cortex; retrograde.
Conflict of interest statement
Conflicts of Interest: The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures















References
-
- Akhter F., Haque T., Sato F., Kato T., Ohara H., Fujio T., Tsutsumi K., Uchino K., Sessle B. J., & Yoshida A. (2014). Projections from the dorsal peduncular cortex to the trigeminal subnucleus caudalis (medullary dorsal horn) and other lower brainstem areas in rats. Neuroscience, 266, 23–37. 10.1016/j.neuroscience.2014.01.046 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous