

Chemogenetic induction of CA1 hyperexcitability triggers indistinguishable autistic traits in asymptomatic mice differing in Ambra1 expression and sex
- PMID: 40097399
- PMCID: PMC11914586
- DOI: 10.1038/s41398-025-03271-0
Chemogenetic induction of CA1 hyperexcitability triggers indistinguishable autistic traits in asymptomatic mice differing in Ambra1 expression and sex
Abstract
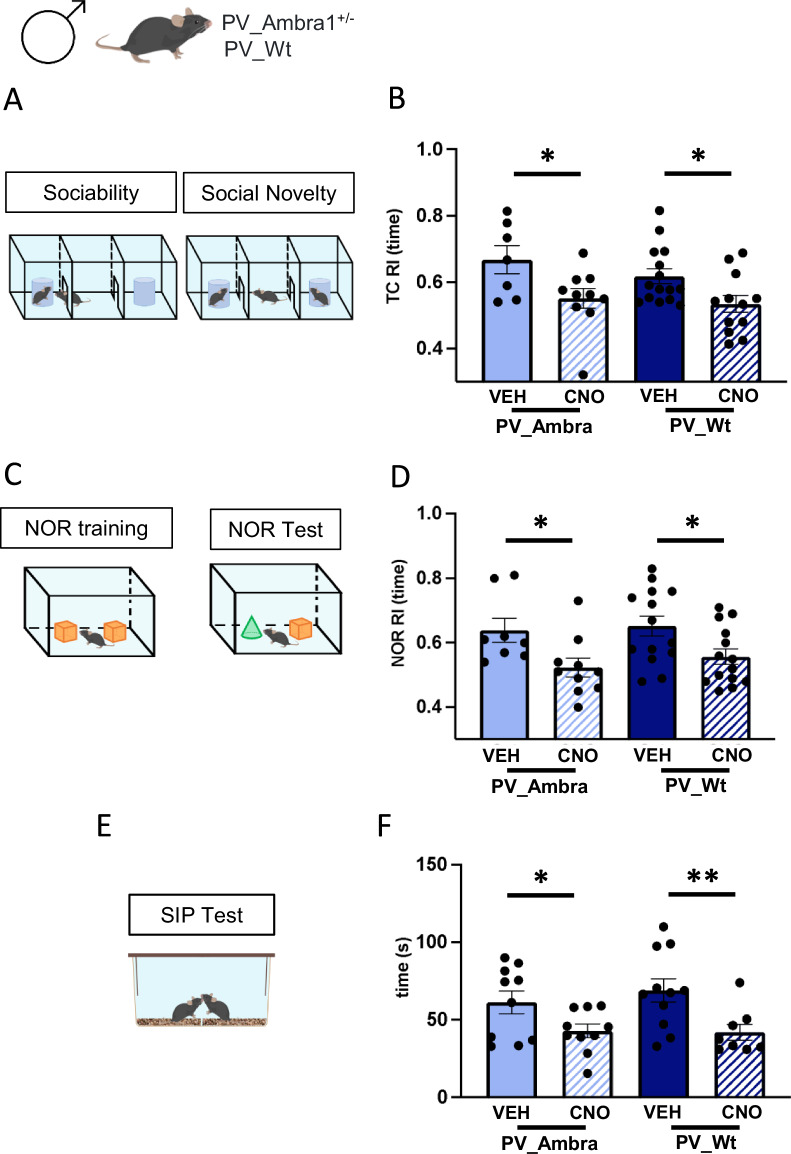
Among the genomic alterations identified as risk factors in mice models of autism spectrum disorders (ASD), heterozygous deletion of Ambra1 (Activating Molecule in Beclin1-Regulated Autophagy) triggers an ASD phenotype associated with hippocampal hyperexcitability exclusively in the female sex although Ambra1 protein is comparably expressed in the hippocampus of symptomatic females and asymptomatic males. Given the intricate relationship between Ambra1 deficiency and sex in the etiology of ASD, we took advantage of asymptomatic mice including Ambra1+/- males and wild-type (Wt) mice of both sexes to investigate whether their non-pathogenic variations in Ambra1 levels could underlie a differential susceptibility to exhibit ASD-like traits in response to experimental elevation of hippocampal excitability. Here we report that selective activation of inhibitory DREADD in CA1 parvalbumin-positive interneurons (PV-IN) reduces GABAergic currents onto pyramidal neurons (PN), causes social and attentional deficits, and augments the proportion of immature/thin spines in CA1 PN dendrites to the same extent in Ambra1+/- males and Wt mice of both sexes. Our findings show that the substantial hippocampal variations in pro-autophagic Ambra1 gene product shown by asymptomatic mice differing in mutation and/or sex do not underlie a differential reactivity to chemogenetic induction of idiopathic ASD.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval: All methods were conducted in strict accordance with relevant guidelines and regulations. Approval for all study-specific experiments and procedures was obtained from the Italian Ministry of Health (Art. 31, D. Lgs 26/2014) based on the ethical guidelines established by the European Council Directive (2010/63/EU). The animal protocol was authorized under permit No. 357/2018-PR.
Figures





References
-
- Ajram LA, Pereira AC, Durieux AMS, Velthius HE, Petrinovic MM, McAlonan GM. The contribution of [1H] magnetic resonance spectroscopy to the study of excitation-inhibition in autism. Prog Neuropsychopharmacol Biol Psychiatry. 2019;89:236–44. - PubMed
-
- Lee E, Lee J, Kim E. Excitation/inhibition imbalance in animal models of autism spectrum disorders. Biol Psychiatry. 2017;81:838–47. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
