

Adipose progenitor cell-derived extracellular vesicles suppress macrophage M1 program to alleviate midlife obesity
- PMID: 40113754
- PMCID: PMC11926339
- DOI: 10.1038/s41467-025-57444-y
Adipose progenitor cell-derived extracellular vesicles suppress macrophage M1 program to alleviate midlife obesity
Abstract
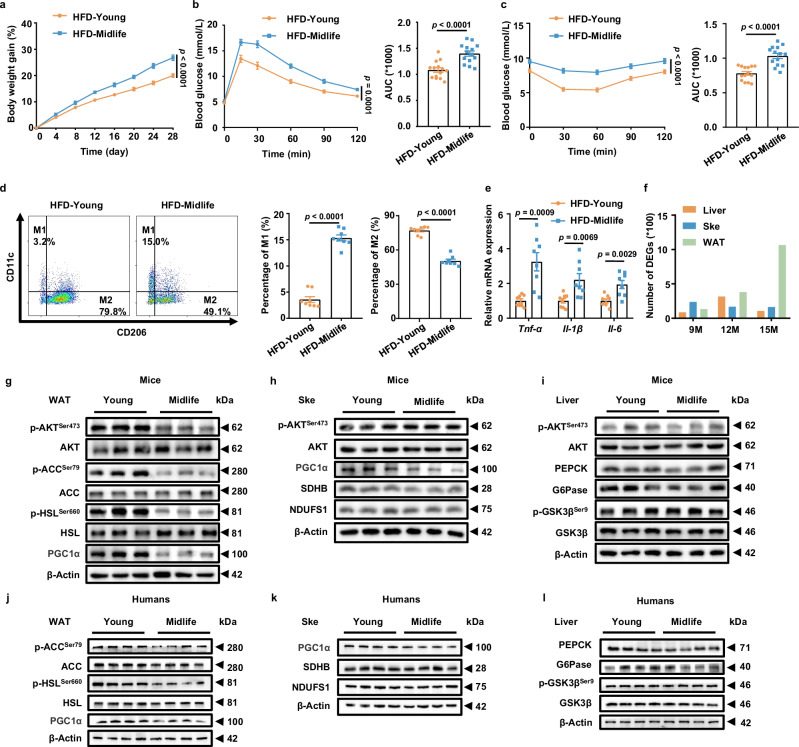
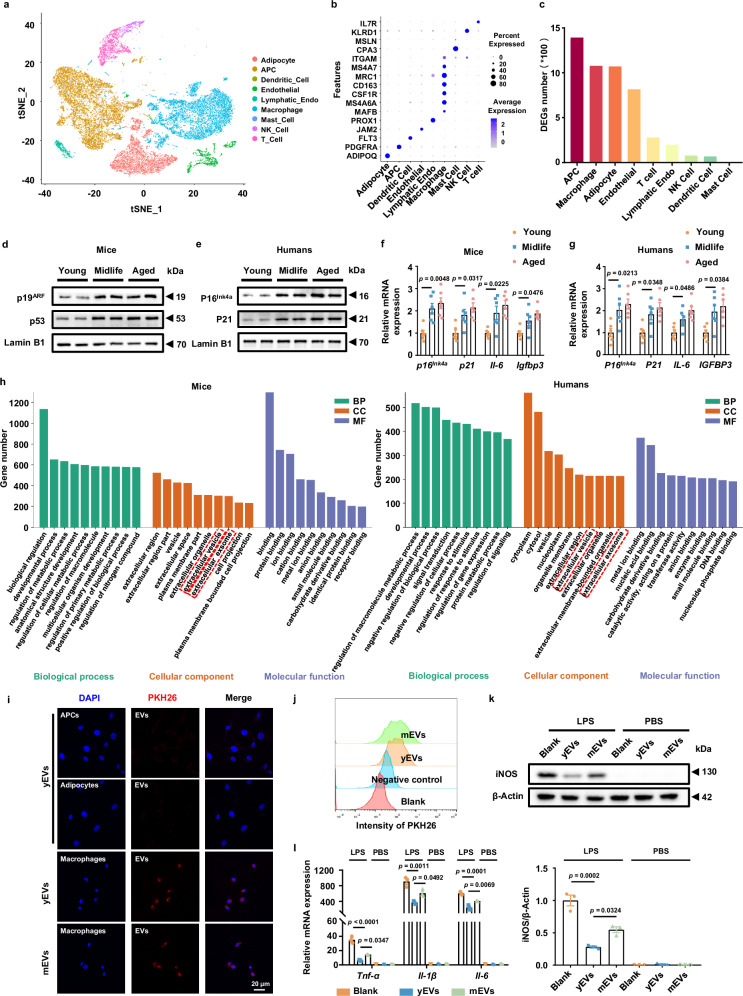
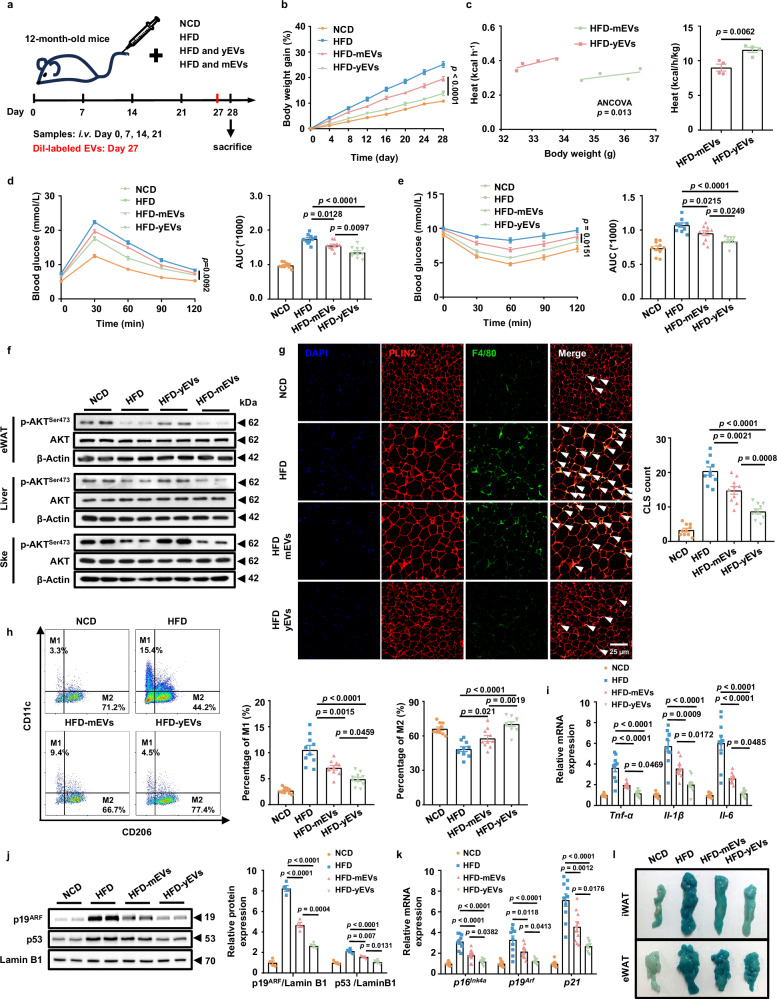
Among different age groups, middle-aged individuals are particularly susceptible to obesity, with a 22% higher risk of all-cause mortality. However, the underlying mechanisms remain unclear. In this study, we identify adipose progenitor cells (APCs) in the white adipose tissue (WAT) of middle-aged subjects as potential causes of midlife obesity. Specifically, the extracellular vesicles (EVs) derived from APCs display an impaired ability to mitigate the inflammaging of adipose tissue macrophages (ATMs) in middle-aged individuals. Mechanistically, these EVs, lacking miR-145-5p, fail to suppress the expression of L-selectin in ATMs, thereby facilitating their M1 program via the NF-κB signaling pathway. In contrast, EVs from young APCs effectively inhibit M1 macrophage polarization. Accordingly, targeted liposomes are designed to deliver miR-145-5p mimics to ATMs, which effectively prevent the obesity in middle-aged mice. Collectively, our findings highlight the role of APC-derived EVs in midlife obesity and propose miR-145-5pas a promising therapeutic target for clinical applications.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
- 82300948/National Natural Science Foundation of China (National Science Foundation of China)
- 82301753/National Natural Science Foundation of China (National Science Foundation of China)
- 82200926/National Natural Science Foundation of China (National Science Foundation of China)
- 82270885/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
