

Peptidylglycine alpha-amidating monooxygenase is important in mice for beta-cell cilia formation and insulin secretion but promotes diabetes risk through beta-cell independent mechanisms
- PMID: 40120979
- PMCID: PMC12090325
- DOI: 10.1016/j.molmet.2025.102123
Peptidylglycine alpha-amidating monooxygenase is important in mice for beta-cell cilia formation and insulin secretion but promotes diabetes risk through beta-cell independent mechanisms
Abstract
Objectives: Carriers of PAM (peptidylglycine alpha-amidating monooxygenase) coding variant alleles have reduced insulinogenic index, higher risk of developing type 2 diabetes (T2D), and islets from heterozygous carriers of the PAM p.Asp563Gly variant display reduced insulin secretion. Exactly how global PAM deficiency contributes to hyperglycemia remains unclear. PAM is the only enzyme capable of converting glycine-extended peptide hormones into amidated products. Like neuropeptide Y (NPY), α-melanocyte stimulating hormone (αMSH), and glucagon-like peptide 1 (GLP-1), islet amyloid polypeptide (IAPP), a beta cell peptide that forms islet amyloid in type 2 diabetes, is a PAM substrate. We hypothesized that Pam deficiency limited to beta cells would lead to reduced insulin secretion, prevent the production of amidated IAPP, and reveal the extent to which loss of Pam in β-cells could accelerate the onset of hyperglycemia in mice.
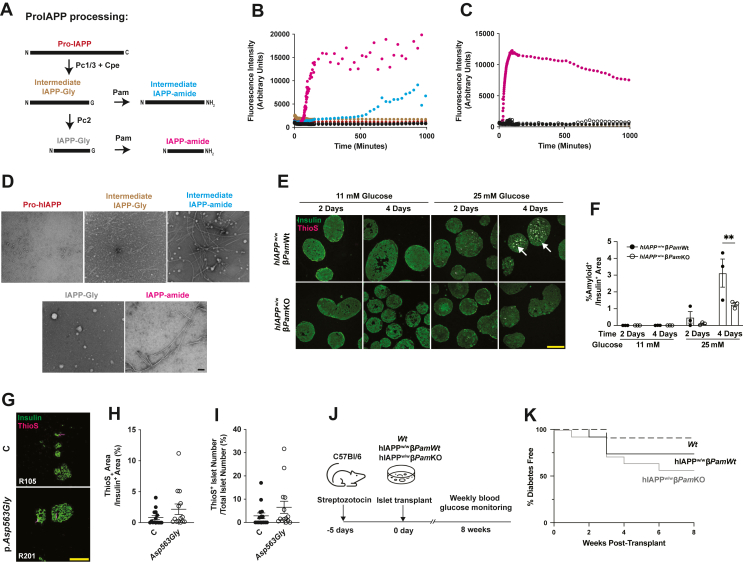
Methods: PAM activity was assessed in human islets from donors based on their PAM genotype. We generated beta cell-specific Pam knockout (Ins1Cre/+, Pamfl/fl; βPamKO) mice and performed islet culture, histological, and metabolic assays to evaluate the physiological roles of Pam in beta cells. We analyzed human IAPP (hIAPP) amyloid fibril forming kinetics using synthetic amidated and non-amidated hIAPP peptides, and generated hIAPP knock-in beta cell-specific Pam knockout (hIAPPw/w βPamKO) mice to determine the impact of hIAPP amidation on islet amyloid burden, islet graft survival, and glucose tolerance.
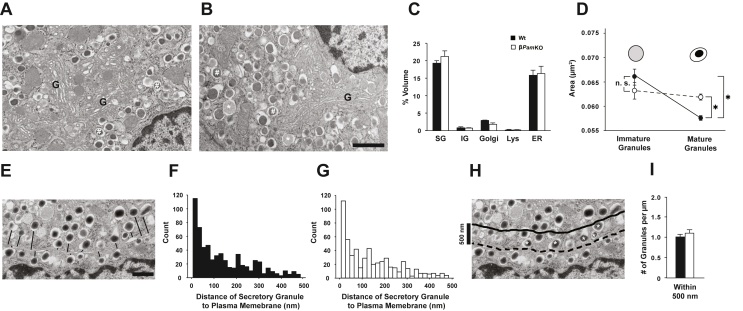
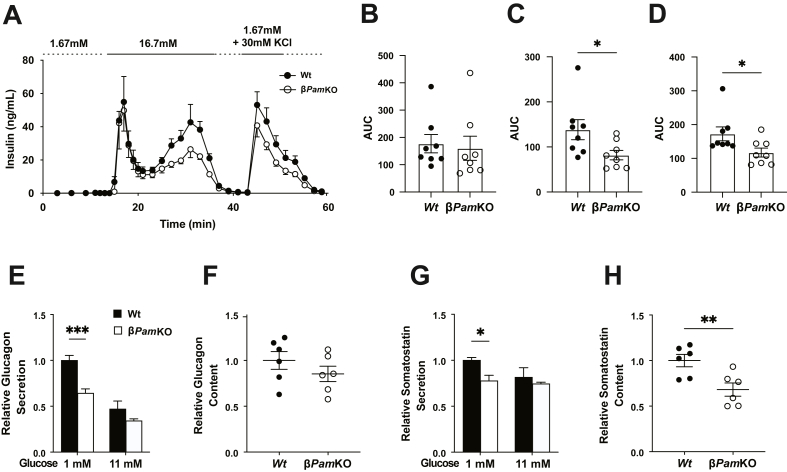
Results: PAM enzyme activity was significantly reduced in islets from donors with the PAM p. Asp563Gly T2D-risk allele. Islets from βPamKO mice had impaired second-phase glucose- and KCl-induced insulin secretion. Beta cells from βPamKO mice had larger dense-core granules and fewer and shorter cilia. Interestingly, non-amidated hIAPP was less fibrillogenic in vitro, and high glucose-treated hIAPPw/w βPamKO islets had reduced amyloid burden. Despite these changes in beta cell function, βPamKO mice were not more susceptible to diet-induced hyperglycemia. In vitro beta cell death and in vivo islet graft survival remained comparable between hIAPPw/w βPamKO and hIAPPw/w islets. Surprisingly, aged hIAPPw/w βPamKO mice had improved insulin secretion and glucose tolerance.
Conclusions: Eliminating Pam expression only in beta cells leads to morphological changes in insulin granules, reduced insulin secretion, reduced hIAPP amyloid burden and altered ciliogenesis. However, in mice beta-cell Pam deficiency has no impact on the development of diet- or hIAPP-induced hyperglycemia. Our data are consistent with current studies revealing ancient, highly conserved roles for peptidergic signaling in the coordination of the diverse signals needed to regulate fundamental processes such as glucose homeostasis.
Keywords: Insulin granules; Islet amyloid polypeptide; Islet cilia; Peptide hormone amidation; Type 2 diabetes risk gene.
Copyright © 2025. Published by Elsevier GmbH.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: A.L.G's spouse is an employee of Genentech and holds stock options in Roche. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
