

Pharmacological targeting of BMAL1 modulates circadian and immune pathways
- PMID: 40133642
- PMCID: PMC12037410
- DOI: 10.1038/s41589-025-01863-x
Pharmacological targeting of BMAL1 modulates circadian and immune pathways
Abstract
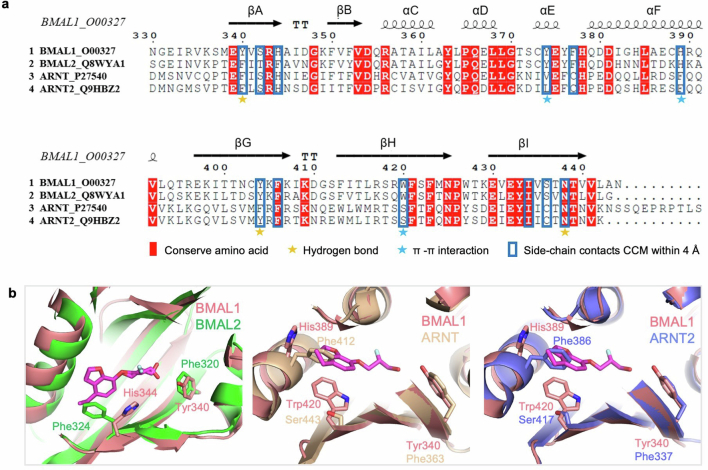
The basic helix-loop-helix PER-ARNT-SIM (bHLH-PAS) proteins BMAL1 and CLOCK heterodimerize to form the master transcription factor governing rhythmic gene expression. Owing to connections between circadian regulation and numerous physiological pathways, targeting the BMAL1-CLOCK complex pharmacologically is an attractive entry point for intervening in circadian-related processes. In this study, we developed a small molecule, Core Circadian Modulator (CCM), that targets the cavity in the PASB domain of BMAL1, causing it to expand, leading to conformational changes in the PASB domain and altering the functions of BMAL1 as a transcription factor. Biochemical, structural and cellular investigations validate the high level of selectivity of CCM in engaging BMAL1, enabling direct access to BMAL1-CLOCK cellular activities. CCM induces dose-dependent alterations in PER2-Luc oscillations and orchestrates the downregulation of inflammatory and phagocytic pathways in macrophages. These findings collectively reveal that the BMAL1 protein architecture is inherently configured to enable the binding of chemical ligands for functional modulation.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: None of the authors has competing financial or non-financial interests as defined by Nature Portfolio.
Figures











References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
