

The clinical antiprotozoal drug halofuginone promotes weight loss by elevating GDF15 and FGF21
- PMID: 40138418
- PMCID: PMC11939056
- DOI: 10.1126/sciadv.adt3142
The clinical antiprotozoal drug halofuginone promotes weight loss by elevating GDF15 and FGF21
Abstract
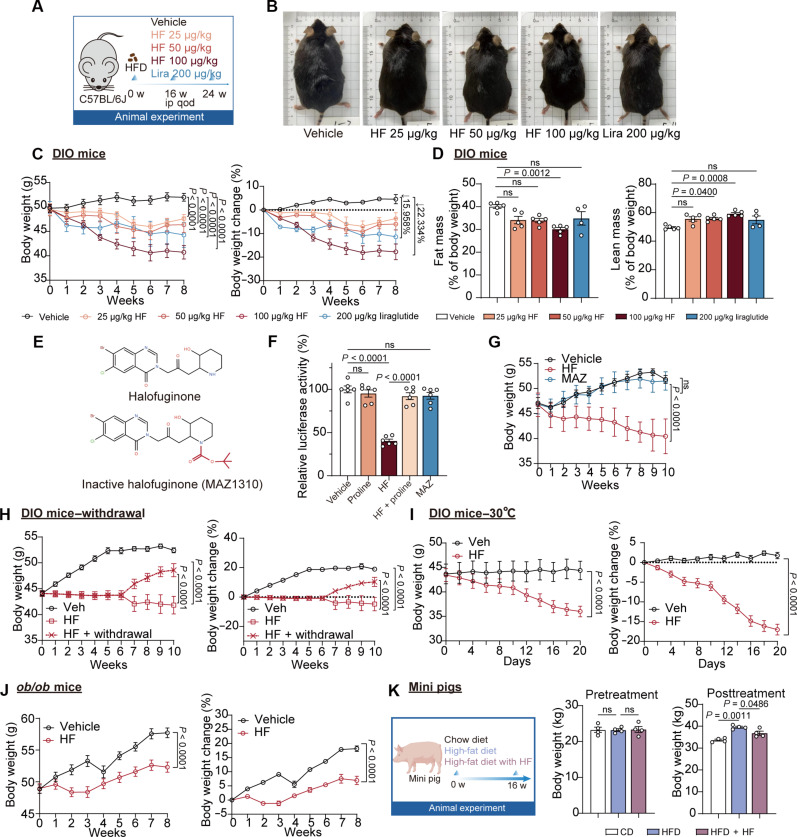
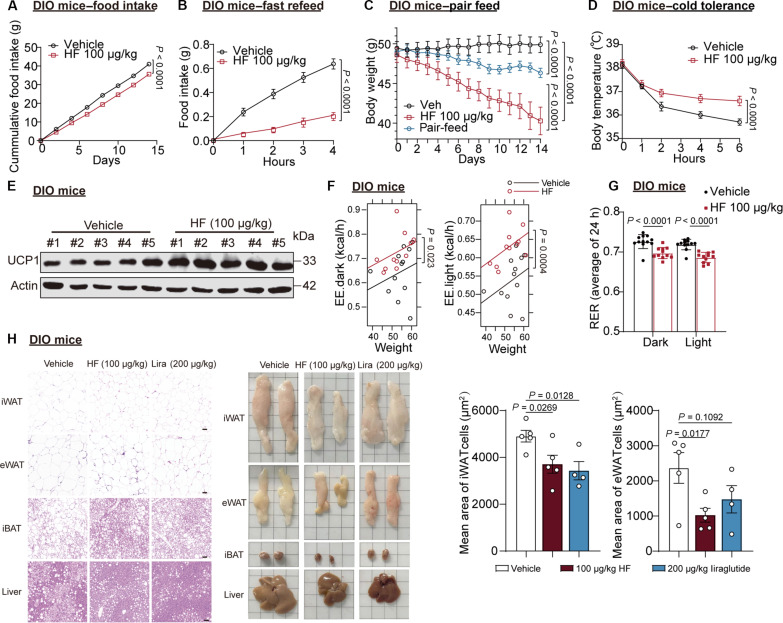
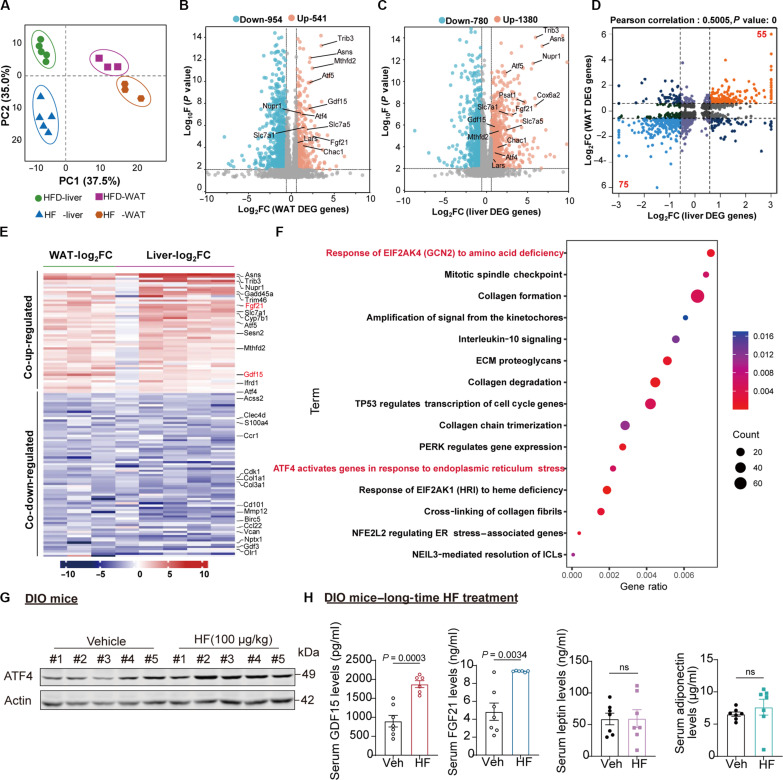
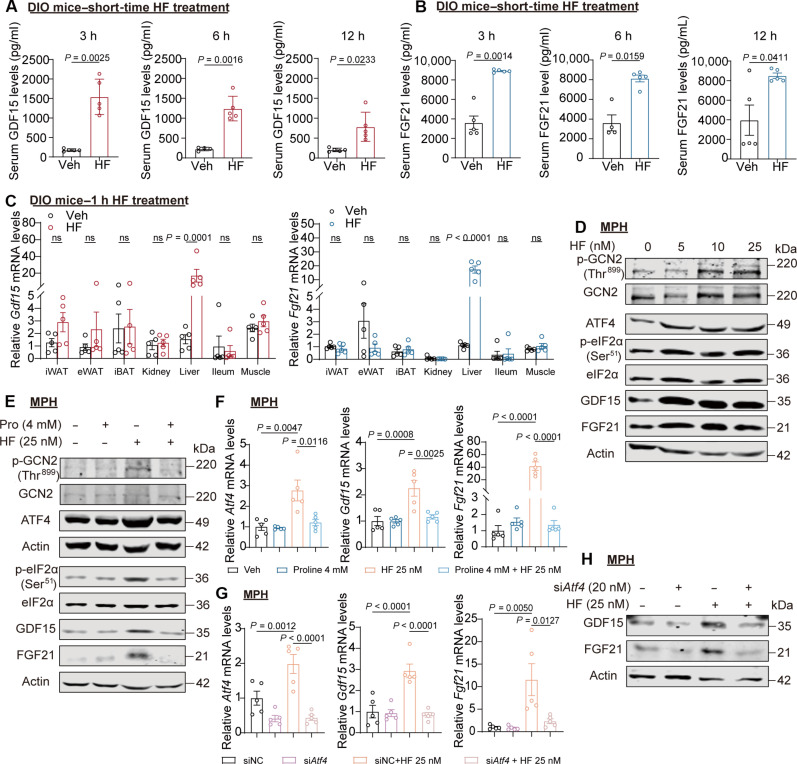
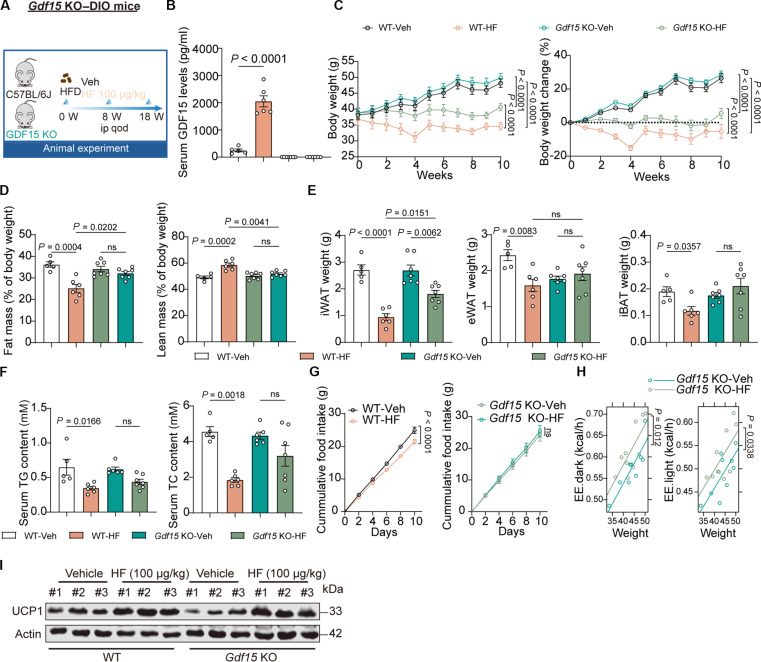
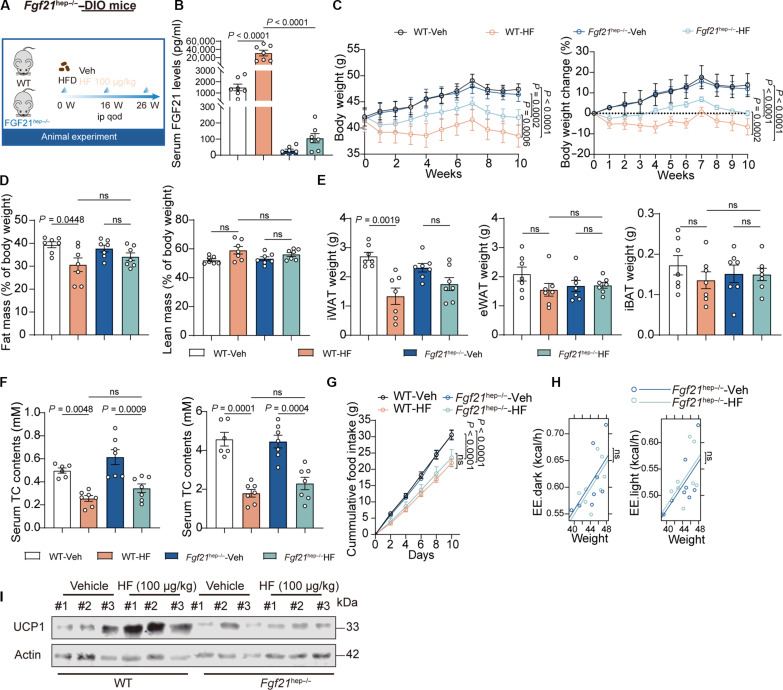
Obesity is a debilitating global pandemic with a huge cost on health care due to it being a major underlying risk factor for several diseases. Therefore, there is an unmet medical need for pharmacological interventions to curb obesity. Here, we report that halofuginone, a Food and Drug Administration-approved anti-scleroderma and antiprotozoal drug, is a promising anti-obesity agent in preclinical mouse and pig models. Halofuginone suppressed food intake, increased energy expenditure, and resulted in weight loss in diet-induced obese mice while also alleviating insulin resistance and hepatic steatosis. Using molecular and pharmacological tools with transcriptomics, we identified that halofuginone increases fibroblast growth factor 21 (FGF21) and growth differentiation factor 15 (GDF15) levels via activating integrated stress response. Using Gdf15 and Fgf21 knockout mice, we show that both hormones are necessary to elicit anti-obesity changes. Together, our study reports the beneficial metabolic effects of halofuginone and underscores its utility in treating obesity and its associated metabolic complications, which merits clinical assessment.
Figures
References
-
- Calle E. E., Rodriguez C., Walker-Thurmond K., Thun M. J., Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med. 348, 1625–1638 (2003). - PubMed
-
- Afshin A., Forouzanfar M. H., Reitsma M. B., Sur P., Estep K., Lee A., Marczak L., Mokdad A. H., Moradi-Lakeh M., Naghavi M., Salama J. S., Vos T., Abate K. H., Abbafati C., Ahmed M. B., Al-Aly Z., Alkerwi A. A., Al-Raddadi R., Amare A. T., Amberbir A., Amegah A. K., Amini E., Amrock S. M., Anjana R. M., Ärnlöv J., Asayesh H., Banerjee A., Barac A., Baye E., Bennett D. A., Beyene A. S., Biadgilign S., Biryukov S., Bjertness E., Boneya D. J., Campos-Nonato I., Carrero J. J., Cecilio P., Cercy K., Ciobanu L. G., Cornaby L., Damtew S. A., Dandona L., Dandona R., Dharmaratne S. D., Duncan B. B., Eshrati B., Esteghamati A., Feigin V. L., Fernandes J. C., Fürst T., Gebrehiwot T. T., Gold A., Gona P. N., Goto A., Habtewold T. D., Hadush K. T., Hafezi-Nejad N., Hay S. I., Horino M., Islami F., Kamal R., Kasaeian A., Katikireddi S. V., Kengne A. P., Kesavachandran C. N., Khader Y. S., Khang Y.-H., Khubchandani J., Kim D., Kim Y. J., Kinfu Y., Kosen S., Ku T., Defo B. K., Kumar G. A., Larson H. J., Leinsalu M., Liang X., Lim S. S., Liu P., Lopez A. D., Lozano R., Majeed A., Malekzadeh R., Malta D. C., Mazidi M., McAlinden C., McGarvey S. T., Mengistu D. T., Mensah G. A., Mensink G. B. M., Mezgebe H. B., Mirrakhimov E. M., Mueller U. O., Noubiap J. J., Obermeyer C. M., Ogbo F. A., Owolabi M. O., Patton G. C., Pourmalek F., Qorbani M., Rafay A., Rai R. K., Ranabhat C. L., Reinig N., Safiri S., Salomon J. A., Sanabria J. R., Santos I. S., Sartorius B., Sawhney M., Schmidhuber J., Schutte A. E., Schmidt M. I., Sepanlou S. G., Shamsizadeh M., Sheikhbahaei S., Shin M.-J., Shiri R., Shiue I., Roba H. S., Silva D. A. S., Silverberg J. I., Singh J. A., Stranges S., Swaminathan S., Tabarés-Seisdedos R., Tadese F., Tedla B. A., Tegegne B. S., Terkawi A. S., Thakur J. S., Tonelli M., Topor-Madry R., Tyrovolas S., Ukwaja K. N., Uthman O. A., Vaezghasemi M., Vasankari T., Vlassov V. V., Vollset S. E., Weiderpass E., Werdecker A., Wesana J., Westerman R., Yano Y., Yonemoto N., Yonga G., Zaidi Z., Zenebe Z. M., Zipkin B., Murray C. J. L., Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 377, 13–27 (2017). - PMC - PubMed
-
- Blüher M., Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 15, 288–298 (2019). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
