

Molecular mechanism of bacteriophage contraction structure of an S-layer-penetrating bacteriophage
- PMID: 40139691
- PMCID: PMC11948020
- DOI: 10.26508/lsa.202403088
Molecular mechanism of bacteriophage contraction structure of an S-layer-penetrating bacteriophage
Abstract
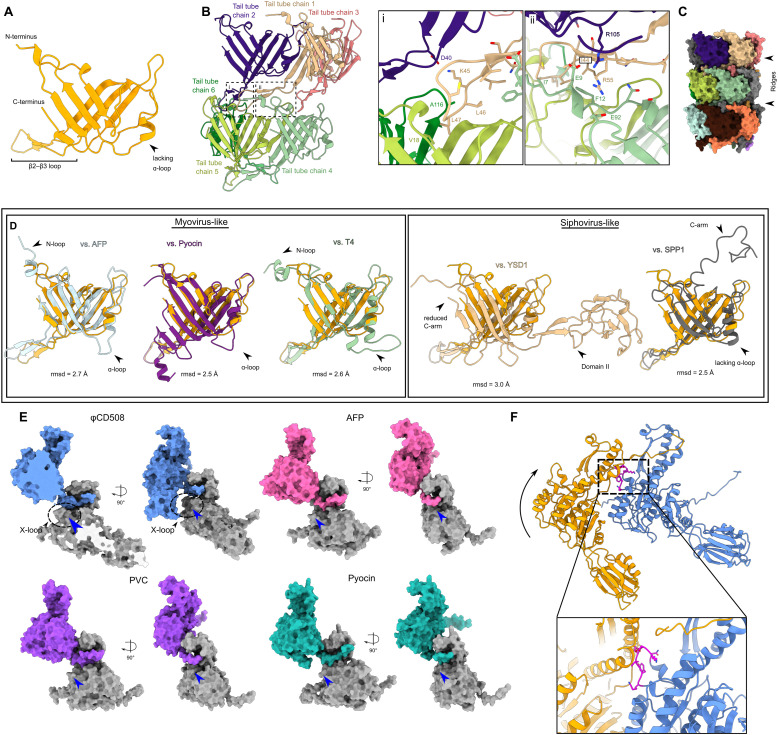
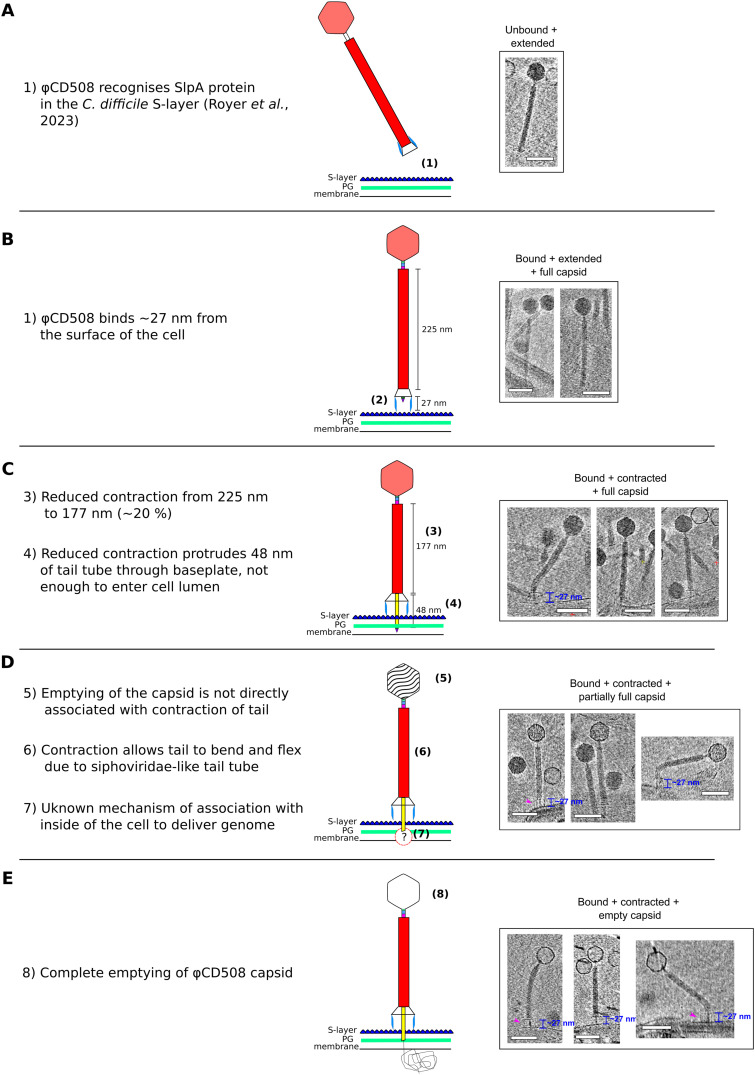
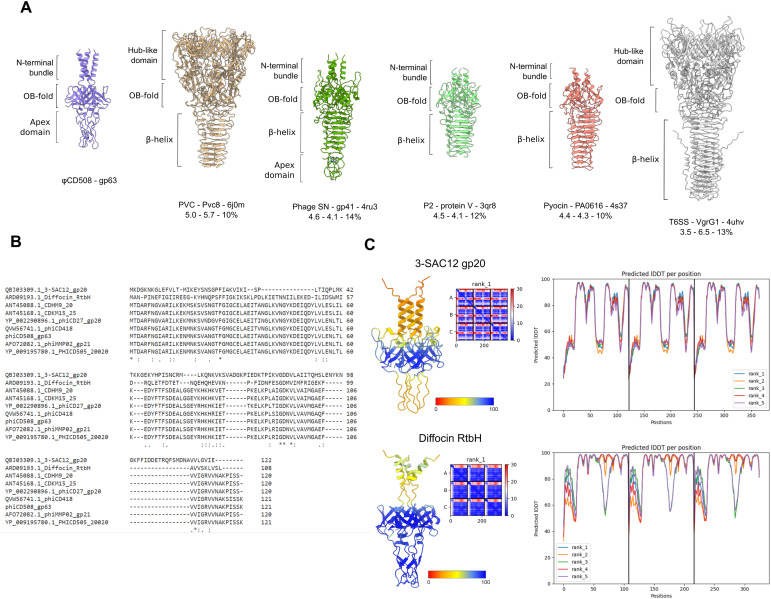
The molecular details of phage tail contraction and bacterial cell envelope penetration remain poorly understood and are completely unknown for phages infecting bacteria enveloped by proteinaceous S-layers. Here, we reveal the extended and contracted atomic structures of an intact contractile-tailed phage (φCD508) that binds to and penetrates the protective S-layer of the Gram-positive human pathogen Clostridioides difficile The tail is unusually long (225 nm), and it is also notable that the tail contracts less than those studied in related contractile injection systems such as the model phage T4 (∼20% compared with ∼50%). Surprisingly, we find no evidence of auxiliary enzymatic domains that other phages exploit in cell wall penetration, suggesting that sufficient energy is released upon tail contraction to penetrate the S-layer and the thick cell wall without enzymatic activity. Instead, the unusually long tail length, which becomes more flexible upon contraction, likely contributes toward the required free energy release for envelope penetration.
© 2025 Wilson et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures












References
-
- Asarnow D, Palovcak E, Cheng Y (2019) asarnow/pyem: UCSF pyem v0.5. Zenodo. Available at: https://zenodo.org/records/3576630.
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources