

The P-loop NTPase RUVBL2 is a conserved clock component across eukaryotes
- PMID: 40140583
- PMCID: PMC12178907
- DOI: 10.1038/s41586-025-08797-3
The P-loop NTPase RUVBL2 is a conserved clock component across eukaryotes
Erratum in
-
Publisher Correction: The P-loop NTPase RUVBL2 is a conserved clock component across eukaryotes.Nature. 2025 Jul;643(8071):E12. doi: 10.1038/s41586-025-09237-y. Nature. 2025. PMID: 40537566 Free PMC article. No abstract available.
Abstract
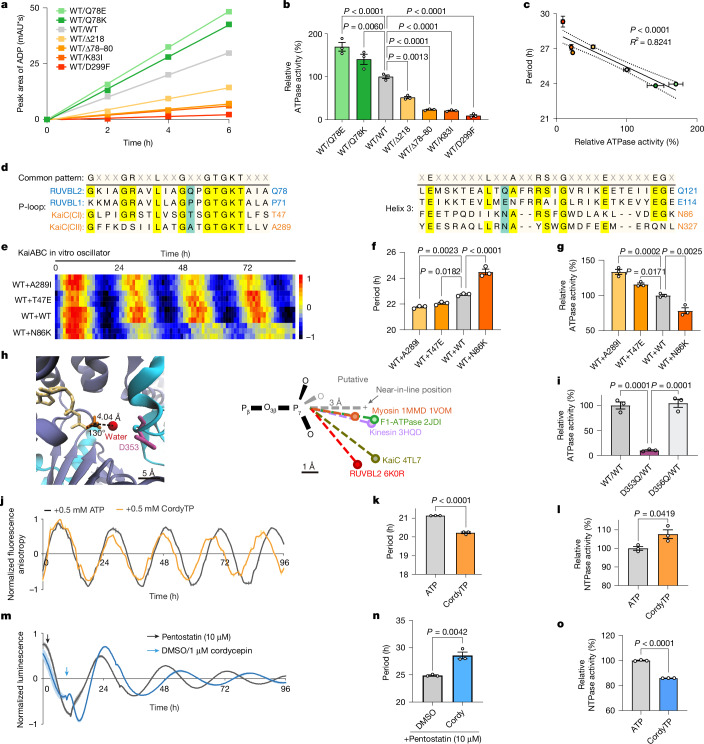
The eukaryotic circadian clock keeps time by using a transcription-translation feedback loop, which exhibits an architecture that is conserved across a diverse range of organisms, including fungi, plants and animals1. Despite their mechanistic similarity, the molecular components of these clocks indicate a lack of common ancestry2. Our study reveals that RUVBL2, which is a P-loop NTPase enzyme previously shown to affect circadian phase and amplitude as part of mammalian clock super-complexes, influences the circadian period through its remarkably slow ATPase activity, resembling the well-characterized KaiC-based clock in cyanobacteria. A screen of RUVBL2 variants identified arrhythmic, short-period and long-period mutants that altered circadian locomotor activity rhythms following delivery by adeno-associated virus to the murine suprachiasmatic nucleus. Enzymatic assays showed that wild-type RUVBL2 hydrolyses only around 13 ATP molecules a day, a vastly reduced turnover compared with typical ATPases. Notably, physical interactions between RUVBL2 orthologues and core clock proteins in humans, Drosophila and the fungus Neurospora, along with consistent circadian phenotypes of RUVBL2-mutant orthologues across species, reinforce their clock-related function in eukaryotes. Thus, as well as establishing RUVBL2 as a common core component in eukaryotic clocks, our study supports the idea that slow ATPase activity, initially discovered in cyanobacteria, is a shared feature of eukaryotic clocks.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: E.E.Z. and D.J. have applied for a patent associated with this study (WIPO publication WO2018133835A1, Nucleoside analog regulating mammalian circadian rhythm).
Figures













References
-
- Zhang, E. E. & Kay, S. A. Clocks not winding down: unravelling circadian networks. Nat. Rev. Mol. Cell Biol.11, 764–776 (2010). - PubMed
-
- Hardin, P. E., Hall, J. C. & Rosbash, M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature343, 536–540 (1990). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
