

Conservation of dichromatin organization along regional centromeres
- PMID: 40147439
- PMCID: PMC12008808
- DOI: 10.1016/j.xgen.2025.100819
Conservation of dichromatin organization along regional centromeres
Abstract
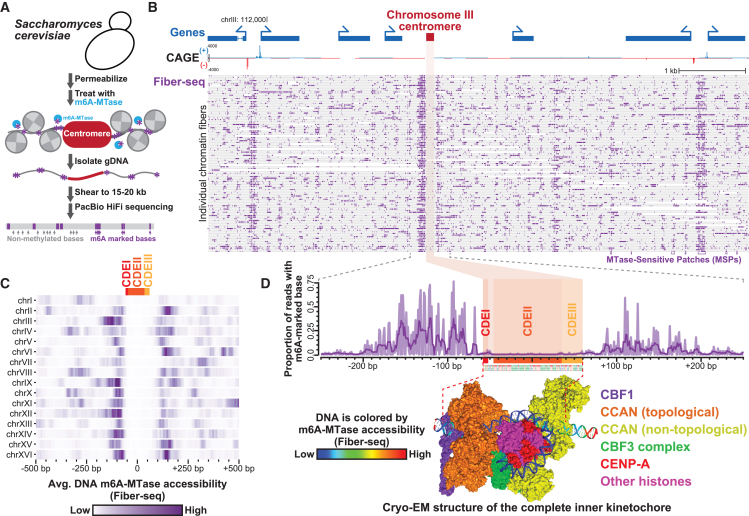
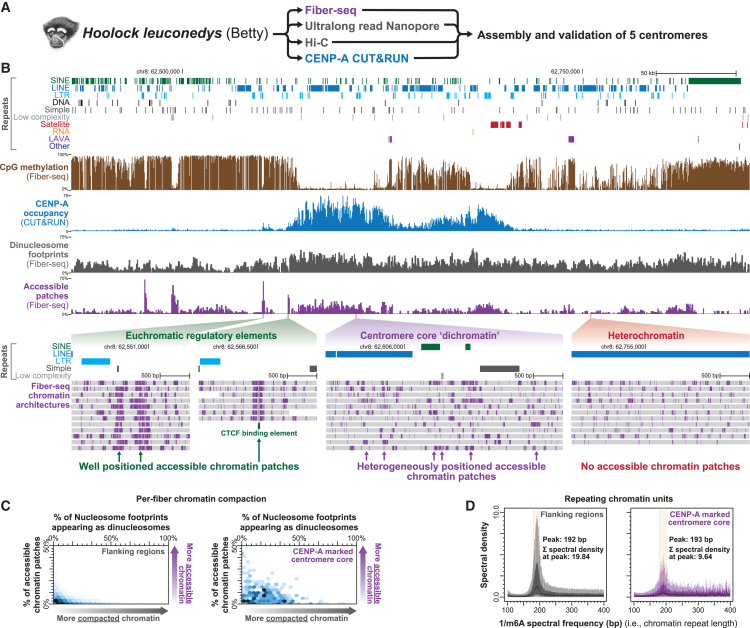
The attachment of the kinetochore to the centromere is essential for genome maintenance, yet the highly repetitive nature of satellite regional centromeres limits our understanding of their chromatin organization. We demonstrate that single-molecule chromatin fiber sequencing (Fiber-seq) can uniquely co-resolve kinetochore and surrounding chromatin architectures along point centromeres, revealing largely homogeneous single-molecule kinetochore occupancy. In contrast, the application of Fiber-seq to regional centromeres exposed marked per-molecule heterogeneity in their chromatin organization. Regional centromere cores uniquely contain a dichotomous chromatin organization (dichromatin) composed of compacted nucleosome arrays punctuated with highly accessible chromatin patches. CENP-B occupancy phases dichromatin to the underlying alpha-satellite repeat within centromere cores but is not necessary for dichromatin formation. Centromere core dichromatin is conserved between humans and primates, including along regional centromeres lacking satellite repeats. Overall, the chromatin organization of regional centromeres is defined by marked per-molecule heterogeneity, buffering kinetochore attachment against sequence and structural variability within regional centromeres.
Keywords: Alpha-satellite; CENP-B; Fiber-seq; centromere; chromatin; kinetochore; single molecule.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.B.S. is a co-inventor on a patent relating to the Fiber-seq method. E.E.E. is a scientific advisory board member of Variant Bio.
Figures





References
-
- Rosenberg H., Singer M., Rosenberg M. Highly reiterated sequences of SIMIANSIMIANSIMIANSIMIANSIMIAN. Science. 1978;200:394–402. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
