

Pathogen-specific stomatal responses in cacao leaves to Phytophthora megakarya and Rhizoctonia solani
- PMID: 40148497
- PMCID: PMC11950177
- DOI: 10.1038/s41598-025-94859-5
Pathogen-specific stomatal responses in cacao leaves to Phytophthora megakarya and Rhizoctonia solani
Abstract
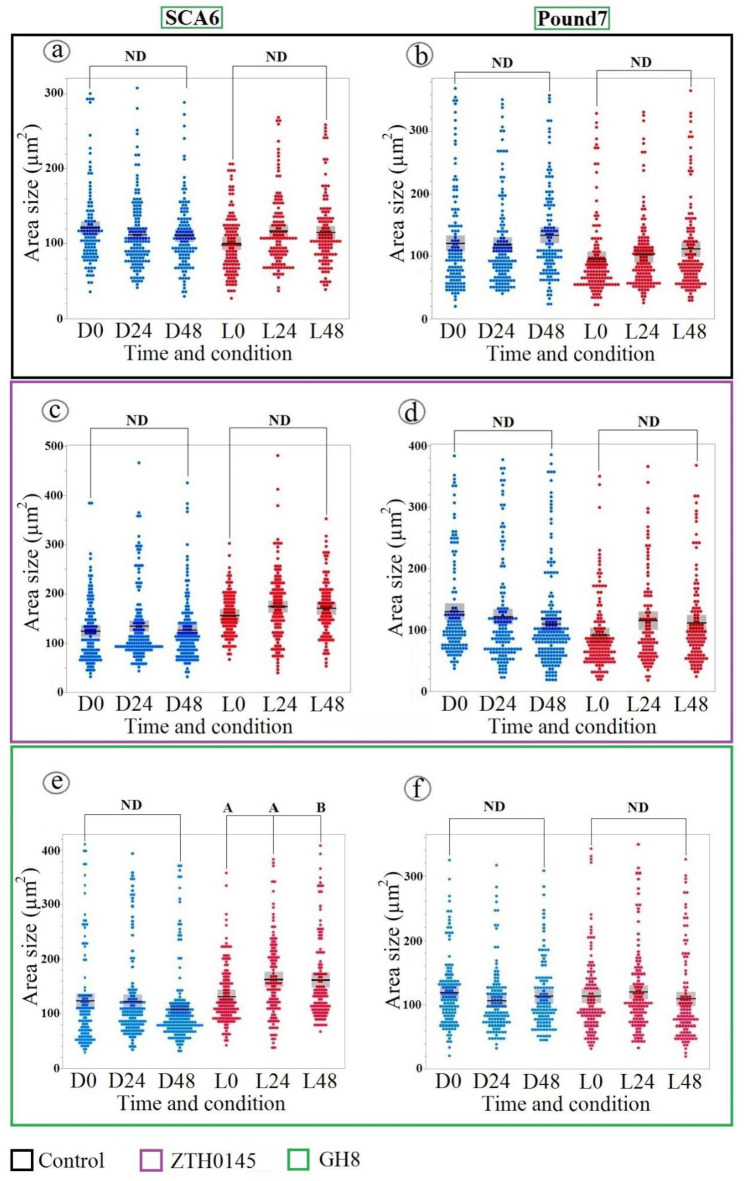
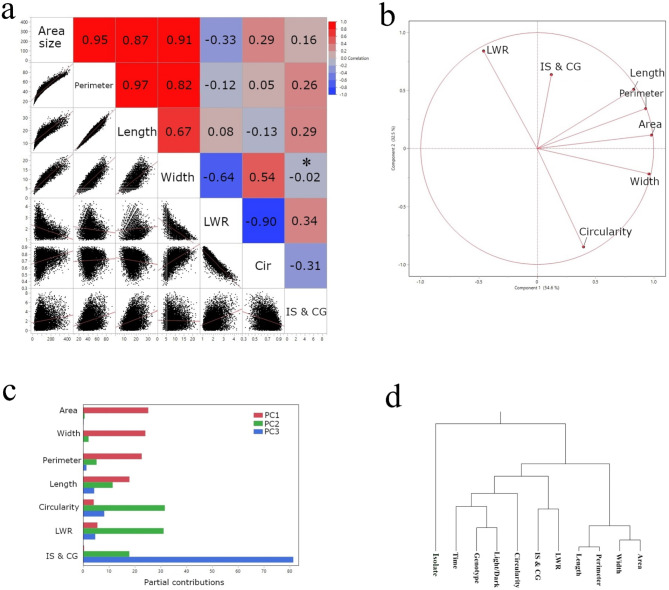
Cacao is a globally significant crop, but its production is severely threatened by diseases, particularly Black Pod Rot (BPR) caused by Phytophthora spp. Understanding plant-pathogen interactions, especially stomatal responses, is crucial for disease management. Machine learning offers a powerful, yet largely untapped, approach to analyze and interpret complex plant responses in plant biology and pathology, particularly in the context of plant-pathogen interactions. This study explores the use of machine learning to analyze and interpret complex stomatal responses in cacao leaves during pathogen interactions. We investigated the impact of the black pod rot pathogen (Phytophthora megakarya) and a non-pathogenic fungus (Rhizoctonia solani) on stomatal aperture in two cacao genotypes (SCA6 and Pound7) under varying light conditions. Image analysis revealed diverse stomatal responses, including no change, opening, and closure, that were influenced by the interplay of genotype, pathogen isolate, and light conditions. Notably, SCA6 exhibited stomatal opening in response to P. megakarya specifically under a 12-hour light/dark cycle, suggesting a light-dependent activation of pathogen virulence factors. In contrast, Pound7 displayed stomatal closure in response to both P. megakarya and R. solani, indicating the potential recognition of conserved Pathogen-Associated Molecular Patterns (PAMPs) and a broader defense response. To further analyze these interactions, we employed machine learning techniques to predict stomatal area size. Our analysis identified key morphological features, with size-related traits being the strongest predictors. Shape-related traits also played a significant role when size-related traits were excluded from the prediction. This study demonstrates the power of combining image analysis and machine learning for discerning subtle, multivariate traits in stomatal dynamics during plant-pathogen interactions, paving the way for future applications in high-throughput disease phenotyping and the development of resistant crop varieties.
Keywords: Black pod rot; Cacao; Light conditions; Machine learning; Stomatal response.
© 2025. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures







References
-
- Vega, C. & Kwik-Uribe, C. Theobroma cacao: An introduction to the plant, its composition, uses, and health benefits. In Cocoa Butter and Related Compounds 35–62 (Elsevier, 2012).
-
- Nieves-Orduña, H. E., Krutovsky, K. V. & Gailing, O. Geographic distribution, conservation, and genomic resources of Cacao theobroma Cacao L. Crop Sci.63, 1750–1778 (2023).
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Miscellaneous
