

Stepwise recruitment of chaperone Hsc70 by DNAJB1 produces ordered arrays primed for bursts of amyloid fibril disassembly
- PMID: 40159506
- PMCID: PMC11955550
- DOI: 10.1038/s42003-025-07906-2
Stepwise recruitment of chaperone Hsc70 by DNAJB1 produces ordered arrays primed for bursts of amyloid fibril disassembly
Abstract
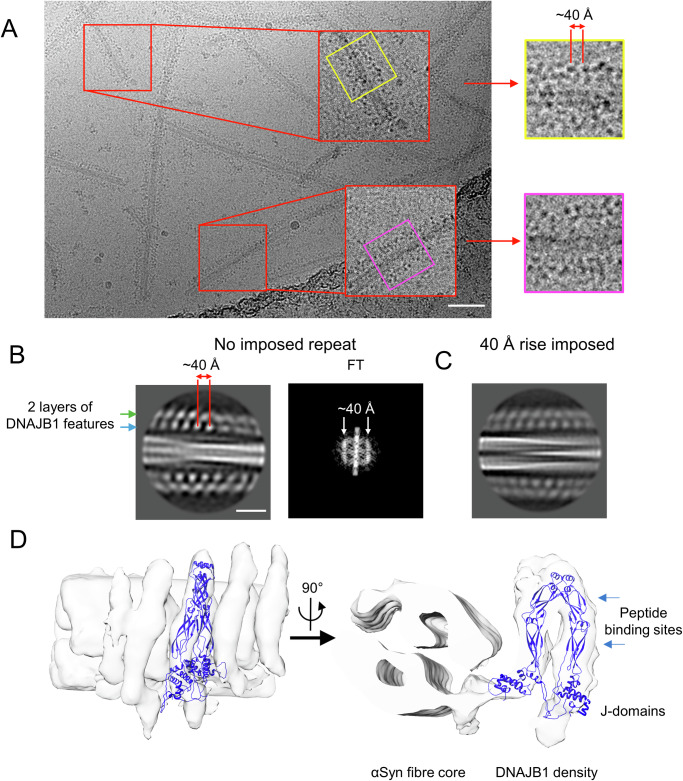
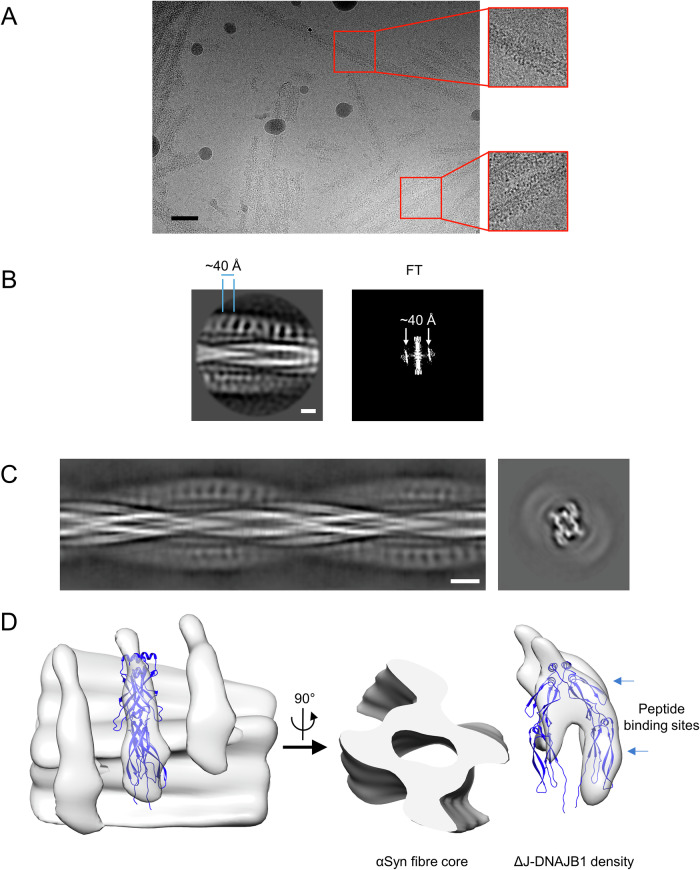
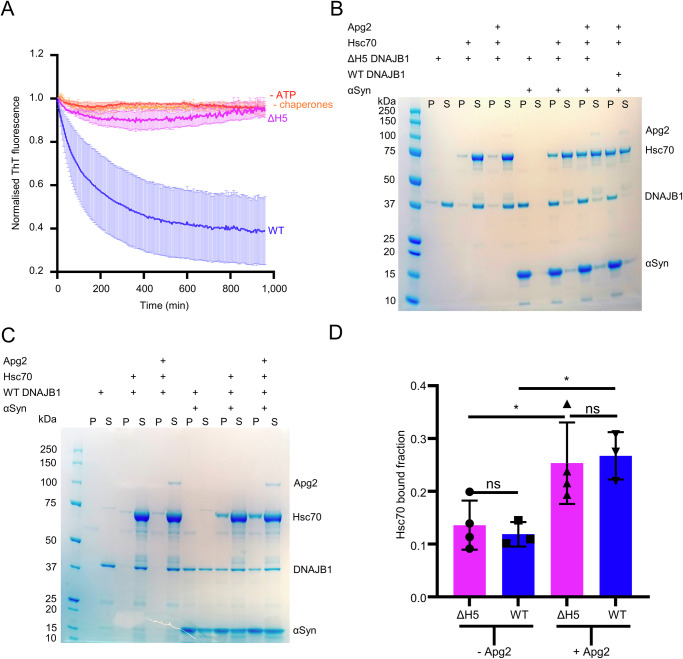
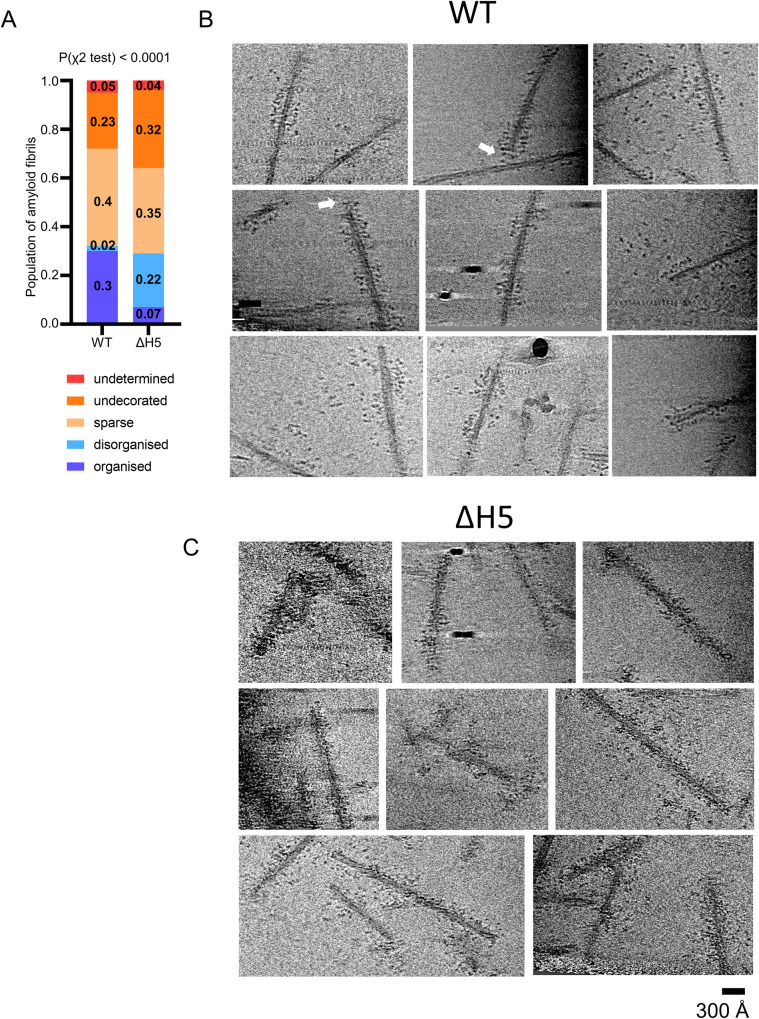
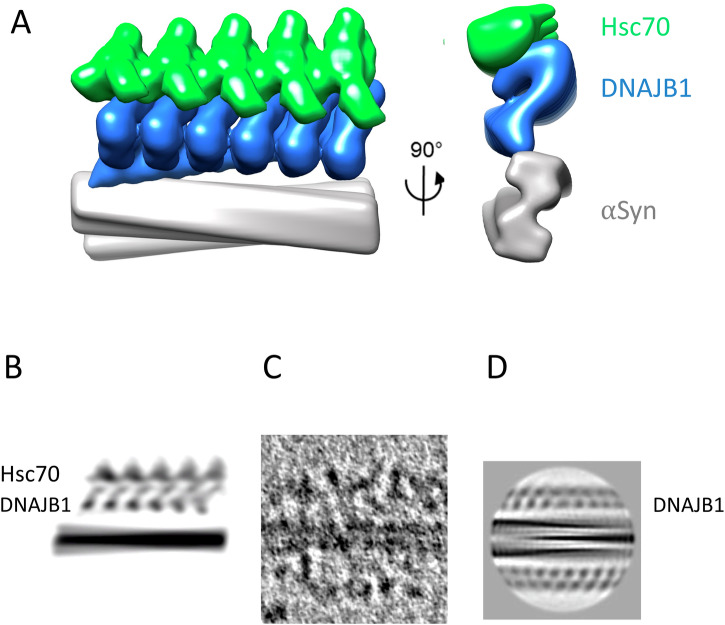
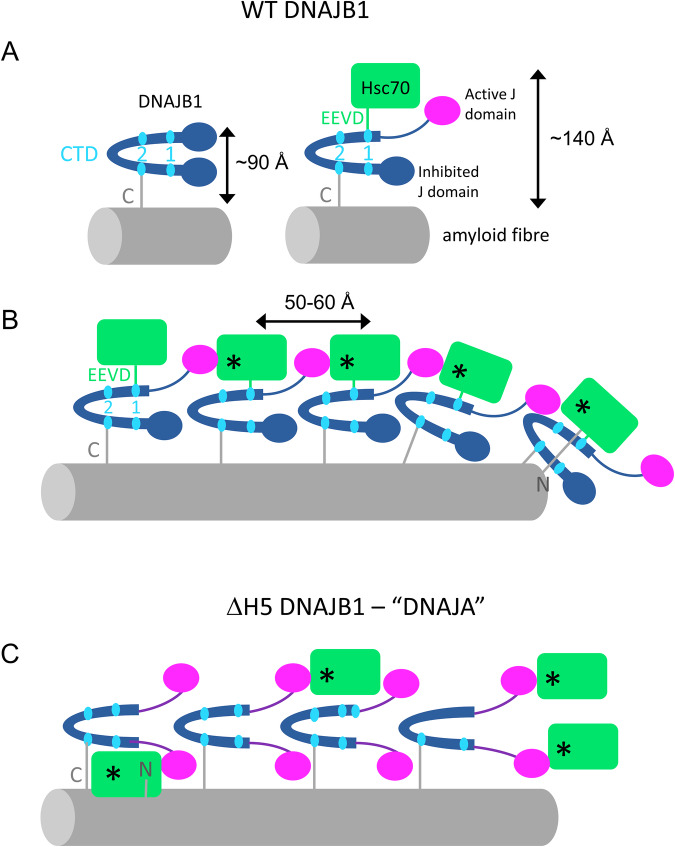
The Hsp70 chaperone system is capable of disassembling pathological aggregates such as amyloid fibres associated with serious degenerative diseases. Here we examine the role of the J-domain protein co-factor in amyloid disaggregation by the Hsc70 system. We used cryo-EM and tomography to compare the assemblies with wild-type DNAJB1 or inactive mutants. We show that DNAJB1 binds regularly along α-synuclein amyloid fibrils and acts in a 2-step recruitment of Hsc70, releasing DNAJB1 auto-inhibition before activating Hsc70 ATPase. The wild-type DNAJB1:Hsc70:Apg2 complex forms dense arrays of chaperones on the fibrils, with Hsc70 on the outer surface. When the auto-inhibition is removed by mutating DNAJB1 (ΔH5 DNAJB1), Hsc70 is recruited to the fibrils at a similar level, but the ΔH5 DNAJB1:Ηsc70:Apg2 complex is inactive, binds less regularly to the fibrils and lacks the ordered clusters. Therefore, we propose that 2-step activation of DNAJB1 regulates the ordered assembly of Hsc70 on the fibril. The localised, dense packing of chaperones could trigger a cascade of recruitment and activation to give coordinated, sequential binding and disaggregation from an exposed fibril end, as previously observed in AFM videos. This mechanism is likely to be important in maintaining a healthy cellular proteome into old age.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

References
-
- Chartier-Harlin, M. et al. α-synuclein locus duplication as a cause of familial Parkinson’s disease. Lancet364, 1169–1171 (2004). - PubMed
-
- Lesage, S. et al. G51D α-synuclein mutation causes a novel Parkinsonian-pyramidal syndrome. Ann. Neurol.73, 459–471 (2013). - PubMed
-
- Polymeropoulos, M. H. et al. Mutation in the α-synuclein gene identified in families with Parkinson’s disease. Science276, 2045–2047 (1997). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
