

This is a preprint.
SORDINO for Silent, Sensitive, Specific, and Artifact-Resisting fMRI in awake behaving mice
- PMID: 40161795
- PMCID: PMC11952411
- DOI: 10.1101/2025.03.10.642406
SORDINO for Silent, Sensitive, Specific, and Artifact-Resisting fMRI in awake behaving mice
Abstract
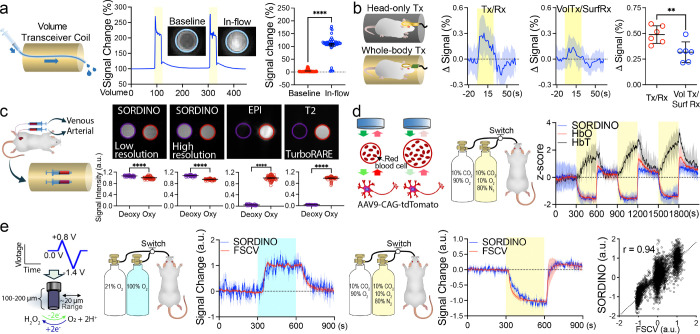
Blood-oxygenation-level-dependent (BOLD) functional magnetic resonance imaging (fMRI) has revolutionized our understanding of the brain activity landscape, bridging circuit neuroscience in animal models with noninvasive brain mapping in humans. This immensely utilized technique, however, faces challenges such as acoustic noise, electromagnetic interference, motion artifacts, magnetic-field inhomogeneity, and limitations in sensitivity and specificity. Here, we introduce Steady-state On-the-Ramp Detection of INduction-decay with Oversampling (SORDINO), a transformative fMRI technique that addresses these challenges by maintaining a constant total gradient amplitude while acquiring data during continuously changing gradient direction. When benchmarked against conventional fMRI on a 9.4T system, SORDINO is silent, sensitive, specific, and resistant to motion and susceptibility artifacts. SORDINO offers superior compatibility with multimodal experiments and carries novel contrast mechanisms distinct from BOLD. It also enables brain-wide activity and connectivity mapping in awake, behaving mice, overcoming stress- and motion-related confounds that are among the most challenging barriers in current animal fMRI studies.
Keywords: T1 contrast; acoustic noise; awake; behavior; brain networks; cerebral blood volume; electromagnetic interference; fMRI; functional connectivity; motion artifacts; mouse; rat; sensitivity; social; specificity; susceptibility artifacts; tissue oxygen.
Conflict of interest statement
Competing Interests Statement The authors declare no competing interests.
Figures





References
Publication types
Grants and funding
- R21 NS133913/NS/NINDS NIH HHS/United States
- R01 EB033790/EB/NIBIB NIH HHS/United States
- RF1 MH117053/MH/NIMH NIH HHS/United States
- R01 NS091236/NS/NINDS NIH HHS/United States
- T32 HD040127/HD/NICHD NIH HHS/United States
- R01 MH126518/MH/NIMH NIH HHS/United States
- S10 MH124745/MH/NIMH NIH HHS/United States
- U01 AA020023/AA/NIAAA NIH HHS/United States
- R01 MH111429/MH/NIMH NIH HHS/United States
- S10 OD026796/OD/NIH HHS/United States
- P60 AA011605/AA/NIAAA NIH HHS/United States
- R21 DA057503/DA/NIDA NIH HHS/United States
- P50 HD103573/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources