

This is a preprint.
Markov models bridge behavioral strategies and circuit principles facilitating thermoregulation
- PMID: 40166317
- PMCID: PMC11957002
- DOI: 10.1101/2025.03.17.643749
Markov models bridge behavioral strategies and circuit principles facilitating thermoregulation
Abstract
Behavioral thermoregulation is critical for survival across animals, including endothermic mammals. However, we do not understand how neural circuits control navigation towards preferred temperatures. Zebrafish exclusively regulate body temperature via behavior, making them ideal for studying thermal navigation. Here, we combine behavioral analysis, machine learning and calcium imaging to understand how larval zebrafish seek out preferred temperatures within thermal gradients. By developing a stimulus-controlled Markov model of thermal navigation we find that hot avoidance largely relies on the modulation of individual swim decisions. The avoidance of cold temperatures, a particular challenge in ectotherms, however relies on a deliberate strategy combining gradient alignment and directed reversals. Calcium imaging identified neurons within the medulla encoding thermal stimuli that form a place-code like representation of the gradient. Our findings establish a key link between neural activity and thermoregulatory behavior, elucidating the neural basis of how animals seek out preferred temperatures.
Conflict of interest statement
Competing Interests The authors declare that no competing interests exist.
Figures
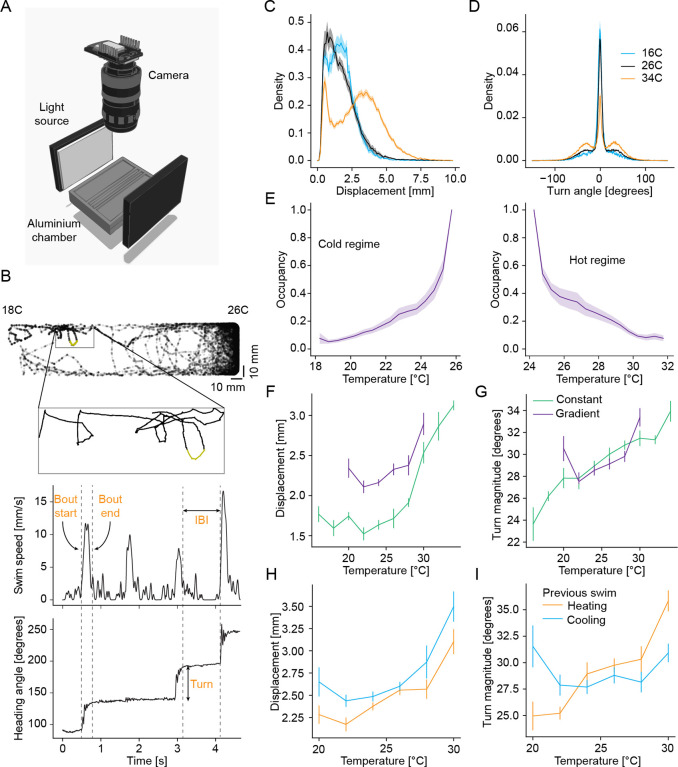

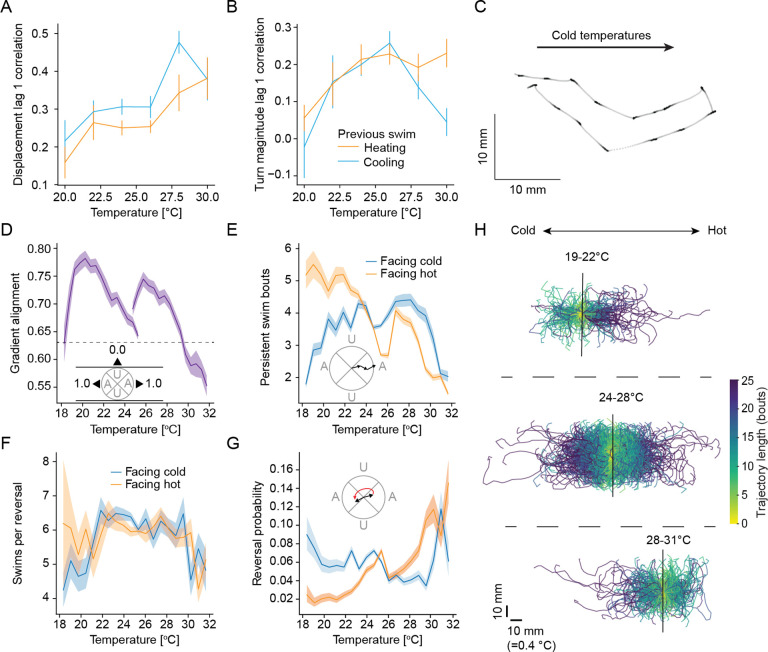



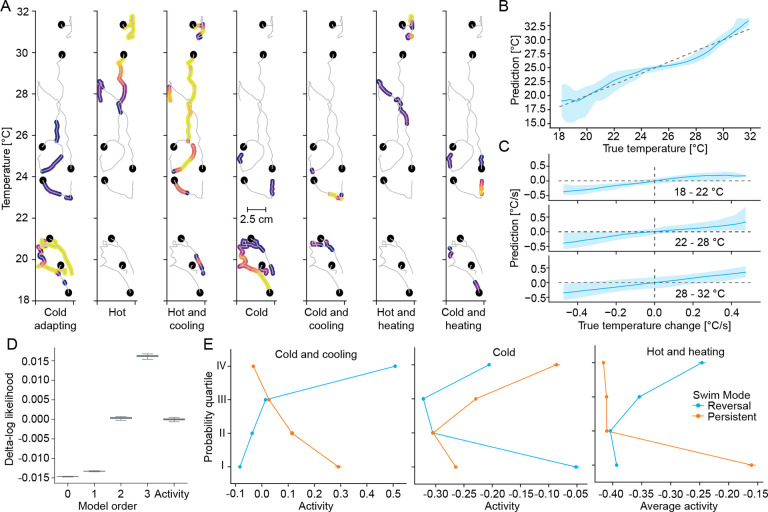
References
-
- Simon H. B. Hyperthermia. N. Engl. J. Med. 329, 483–487 (1993). - PubMed
-
- Matsui T., Ishikawa T., Takeuchi H., Tsukahara M. & Maekawa T. Mild hypothermia inhibits IL-10 production in peripheral blood mononuclear cells. Acta Anaesthesiol. Scand. 48, 205–210 (2004). - PubMed
-
- Wood T. & Thoresen M. Physiological responses to hypothermia. Semin. Fetal Neonatal Med. 20, 87–96 (2015). - PubMed
-
- Bogert C. M. Thermoregulation in reptiles, a factor in evolution. Evolution 3, 195–211 (1949). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources