

Lipotoxicity Induces β-cell Small Extracellular Vesicle-Mediated β-cell Dysfunction in Male Mice
- PMID: 40179251
- PMCID: PMC12006739
- DOI: 10.1210/endocr/bqaf067
Lipotoxicity Induces β-cell Small Extracellular Vesicle-Mediated β-cell Dysfunction in Male Mice
Abstract
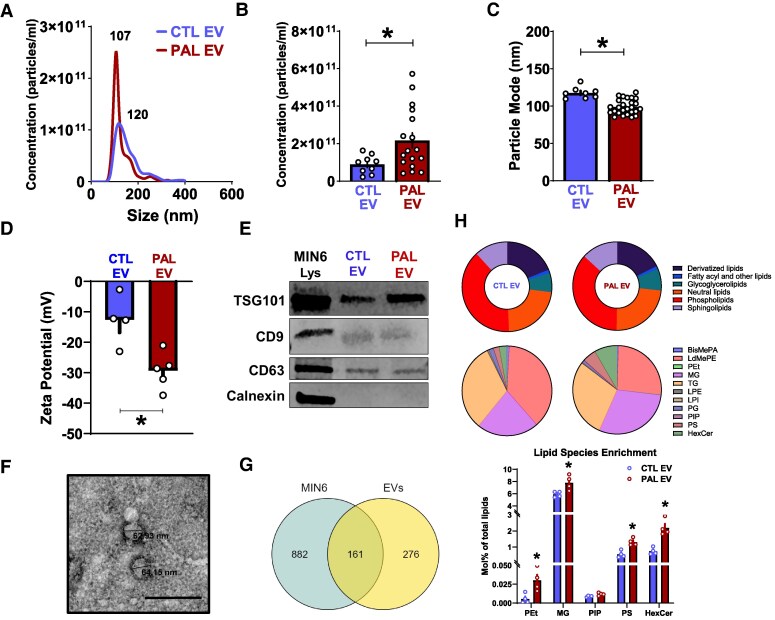
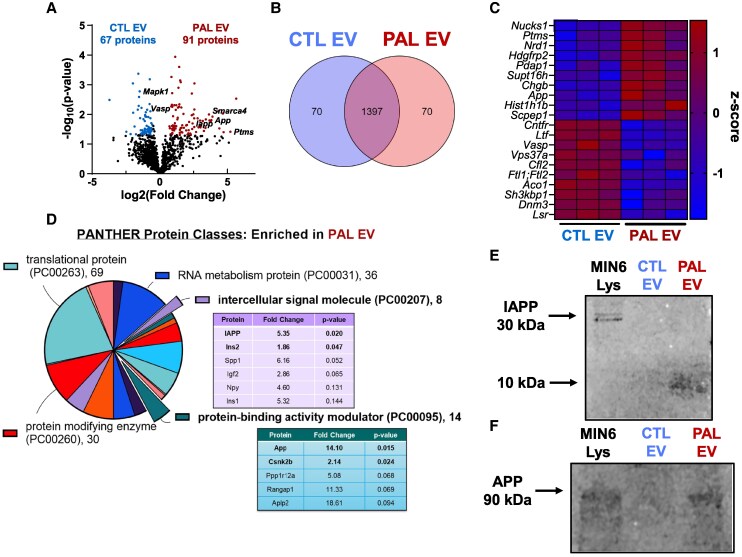
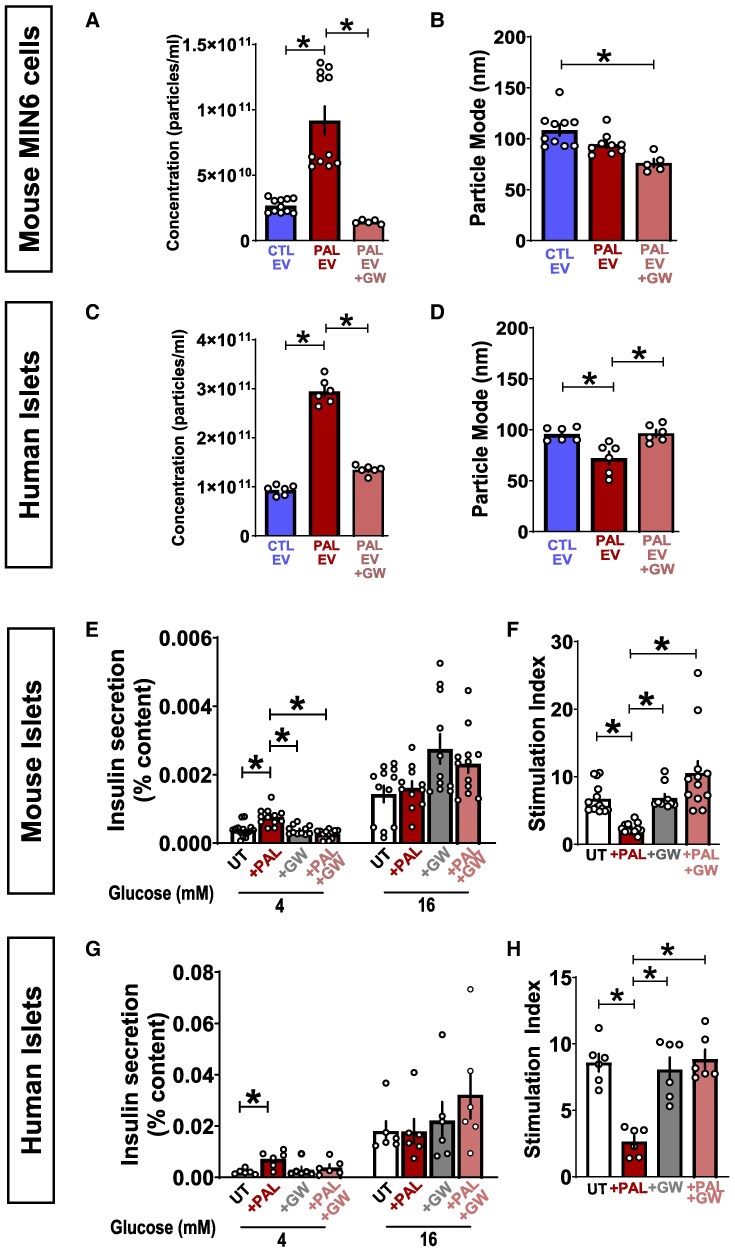
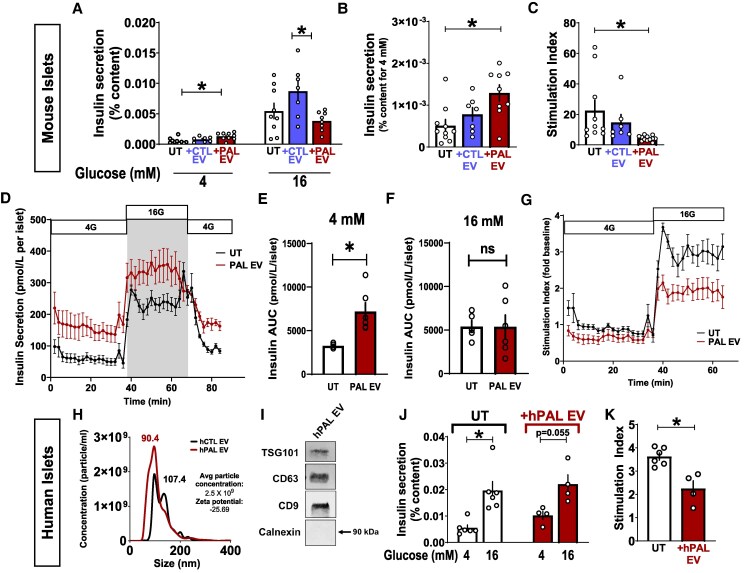
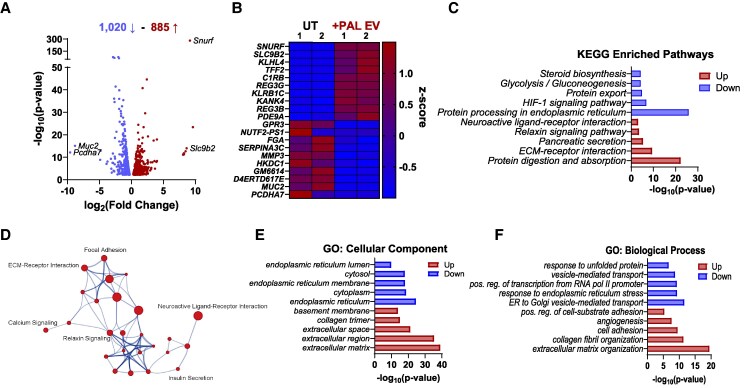
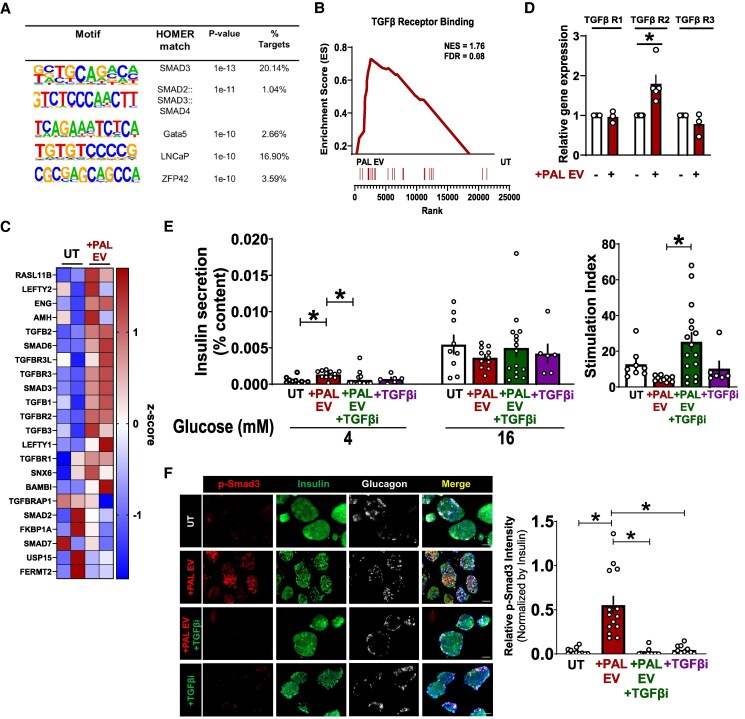
Chronically elevated circulating excess free fatty acids (ie, lipotoxicity) is a pathological process implicated in several metabolic disorders, including obesity-driven type 2 diabetes (T2D). Lipotoxicity exerts detrimental effects on pancreatic islet β-cells by reducing glucose-stimulated insulin secretion (GSIS), altering β-cell transcriptional identity, and promoting apoptosis. While β-cell-derived small extracellular vesicles (sEV) have been shown to contribute to β-cell failure in T2D, their specific role in lipotoxicity-mediated β-cell failure remains to be elucidated. In this work, we demonstrate that lipotoxicity enhances the release of sEVs from β-cells, which exhibit altered proteomic and lipidomic profiles. These palmitate (PAL)-exposed extracellular vesicles (EVs) induce β-cell dysfunction in healthy mouse and human islets and trigger significant islet transcriptional changes, including the upregulation of genes associated with the TGFβ/Smad3 pathway, as noted by RNA sequencing. Importantly, pharmacological inhibition of the TGFβI/II receptor improved PAL EV-induced β-cell dysfunction, underscoring their involvement in activating the TGFβ/Smad3 pathway during this process. We have comprehensively characterized lipotoxic β-cell sEVs and implicated their role in inducing β-cell functional failure in T2D. These findings highlight potential avenues for therapeutic interventions targeting sEV-mediated pathways to preserve β-cell health in metabolic disorders.
Keywords: diabetes; extracellular vesicles; lipotoxicity; β-cell.
© The Author(s) 2025. Published by Oxford University Press on behalf of the Endocrine Society.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
