

Changes in adrenoceptor expression level contribute to the cellular plasticity of glioblastoma cells
- PMID: 40184918
- PMCID: PMC12002996
- DOI: 10.1016/j.jphyss.2025.100016
Changes in adrenoceptor expression level contribute to the cellular plasticity of glioblastoma cells
Abstract
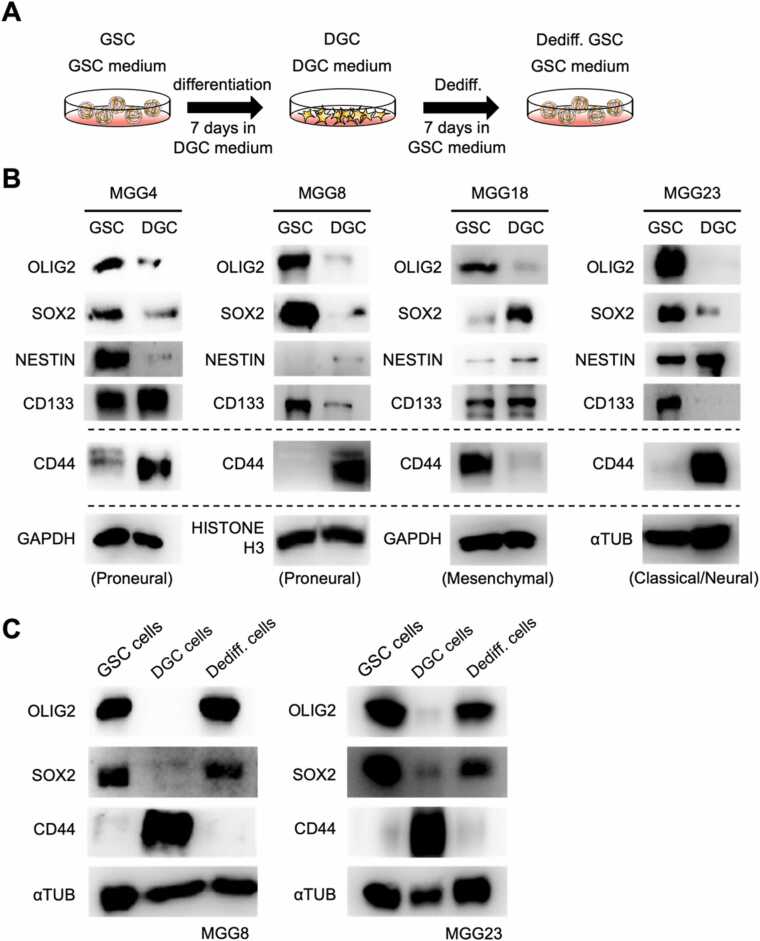
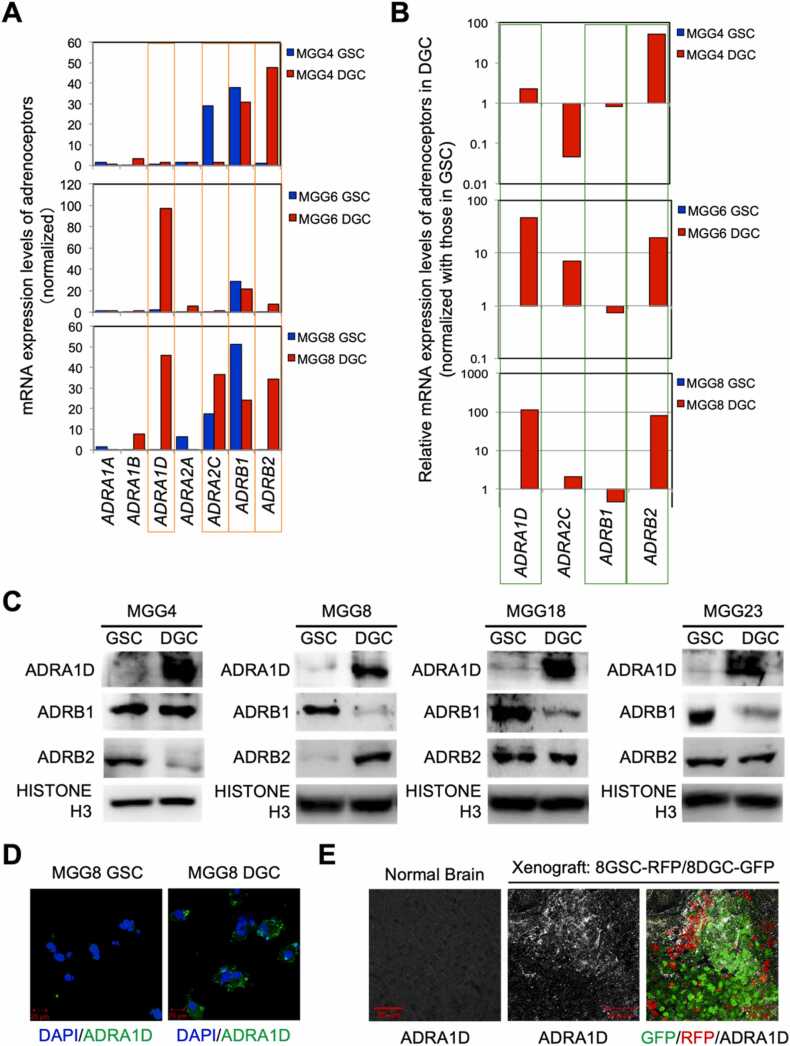
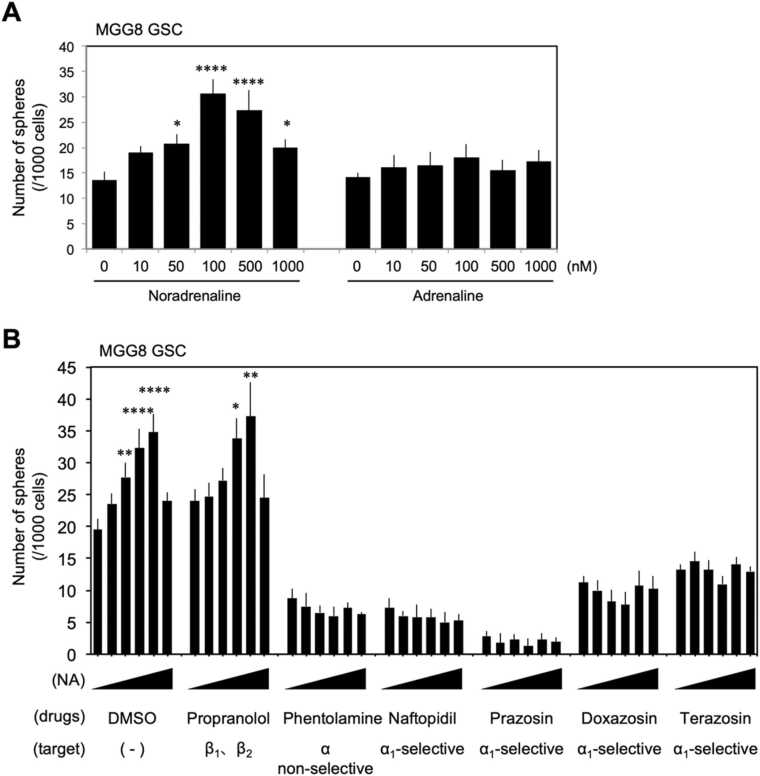
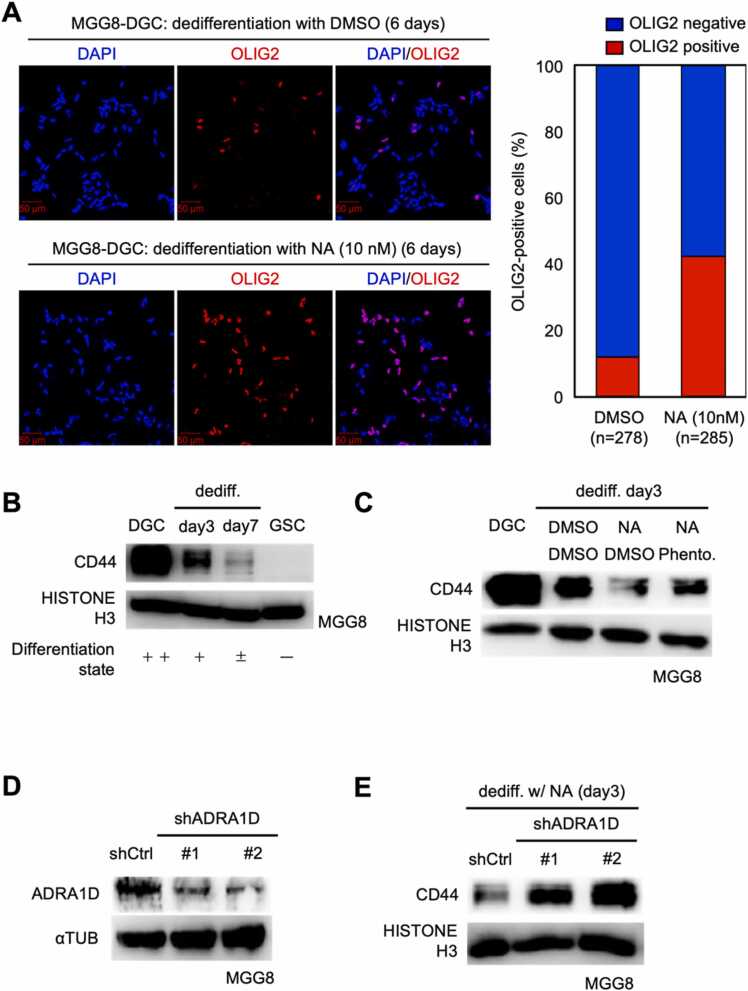
Glioblastoma cells are known to regulate their cellular plasticity in response to their surrounding microenvironment, but it is not fully understood what factors contribute to the cells' changing plasticity. Here, we found that glioblastoma cells alter the expression level of adrenoreceptors depending on their differentiation stage. Catecholamines are abundant in the central nervous system, and we found that noradrenaline, in particular, enhances the stemness of glioblastoma cells and promotes the dedifferentiation potential of already differentiated glioblastoma cells. Antagonist and RNAi experiments revealed that signaling through α1D-adrenoreceptor is important for noradrenaline action on glioblastoma cells. We also found that high α1D-adrenoreceptor expression was associated with poor prognosis in patients with gliomas. These data suggest that glioblastoma cells increase the expression level of their own adrenoreceptors to alter the surrounding tumor microenvironment favorably for survival. We believe that our findings will contribute to the development of new therapeutic strategies for glioblastoma.
Keywords: Adrenoceptors; Cellular plasticity; Differentiated glioma cells; Glioma stem-like cells; Noradrenaline.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures


References
-
- Bao S., Wu Q., McLendon R.E., Hao Y., Shi Q., Hjelmeland A.B., et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444(7120):756–760. Dec 7. - PubMed
-
- Berg T.J., Marques C., Pantazopoulou V., Johansson E., von Stedingk K., Lindgren D., et al. The irradiated brain microenvironment supports glioma stemness and survival via astrocyte-derived transglutaminase 2. Cancer Res. 2021;81(8):2101–2115. Apr 15. - PubMed
-
- Caja L., Bellomo C., Moustakas A. Transforming growth factor β and bone morphogenetic protein actions in brain tumors. FEBS Lett. 2015;589(14):1588–1597. Jun 22. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
