

This is a preprint.
Androgen Deprivation-Induced TET2 Activation Fuels Prostate Cancer Progression via Epigenetic Priming and Slow-Cycling Cancer Cells
- PMID: 40196510
- PMCID: PMC11974783
- DOI: 10.1101/2025.03.26.645495
Androgen Deprivation-Induced TET2 Activation Fuels Prostate Cancer Progression via Epigenetic Priming and Slow-Cycling Cancer Cells
Abstract
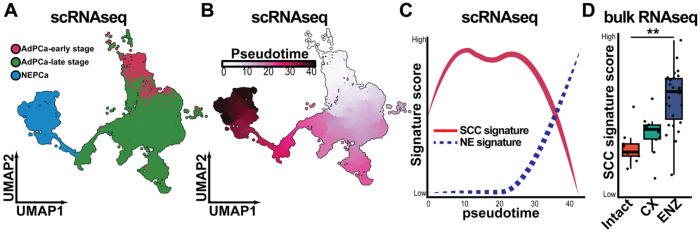
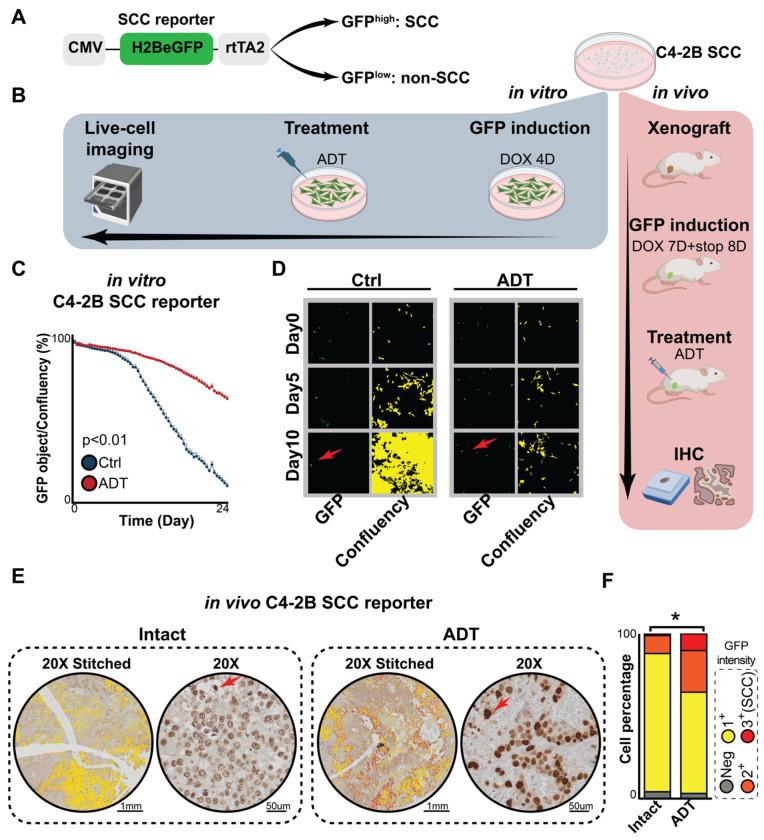
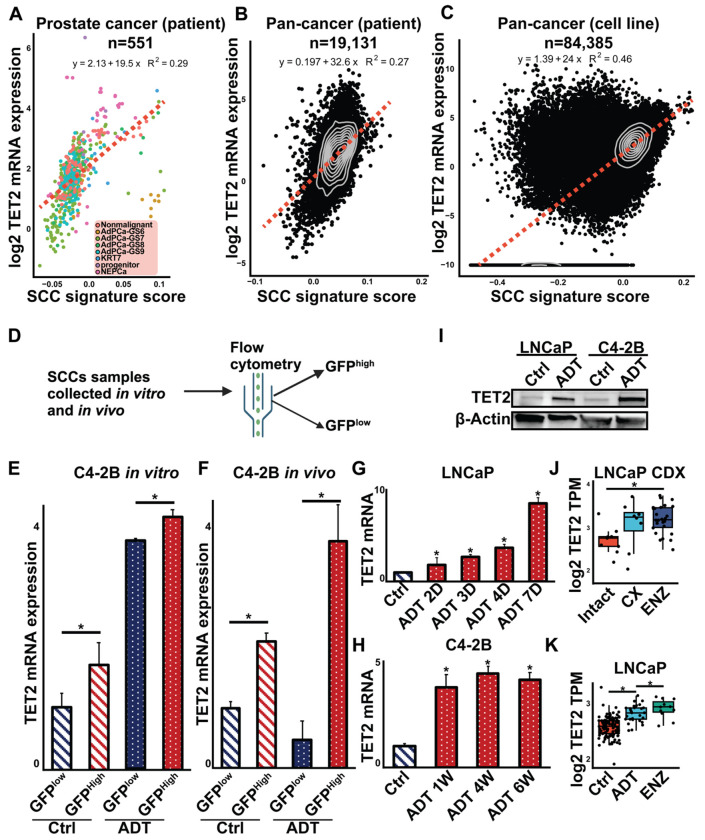
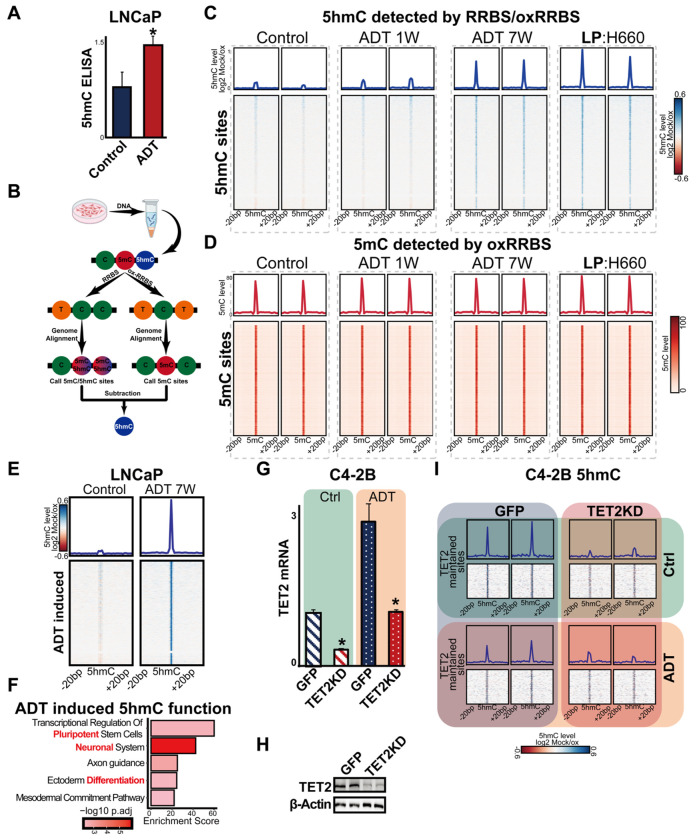
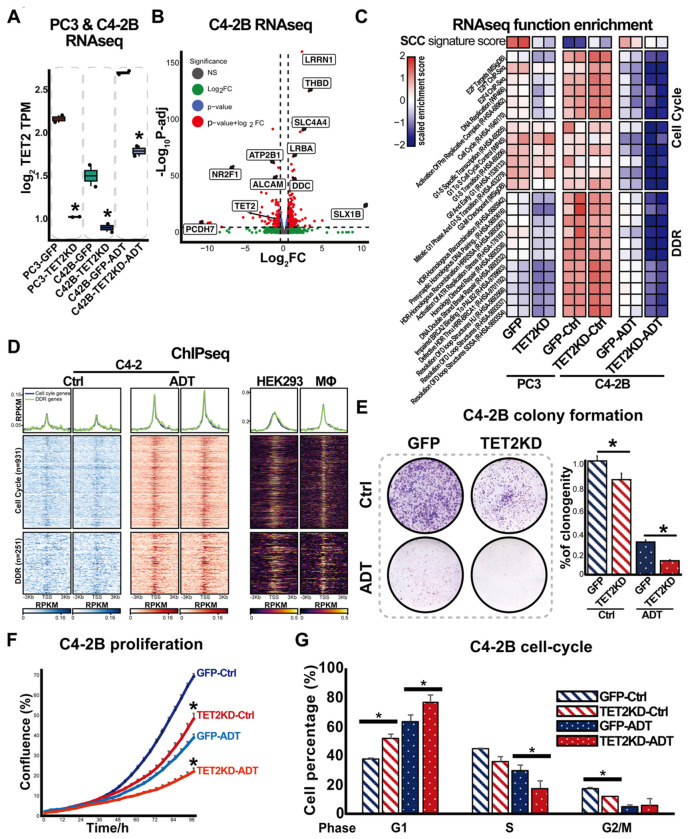
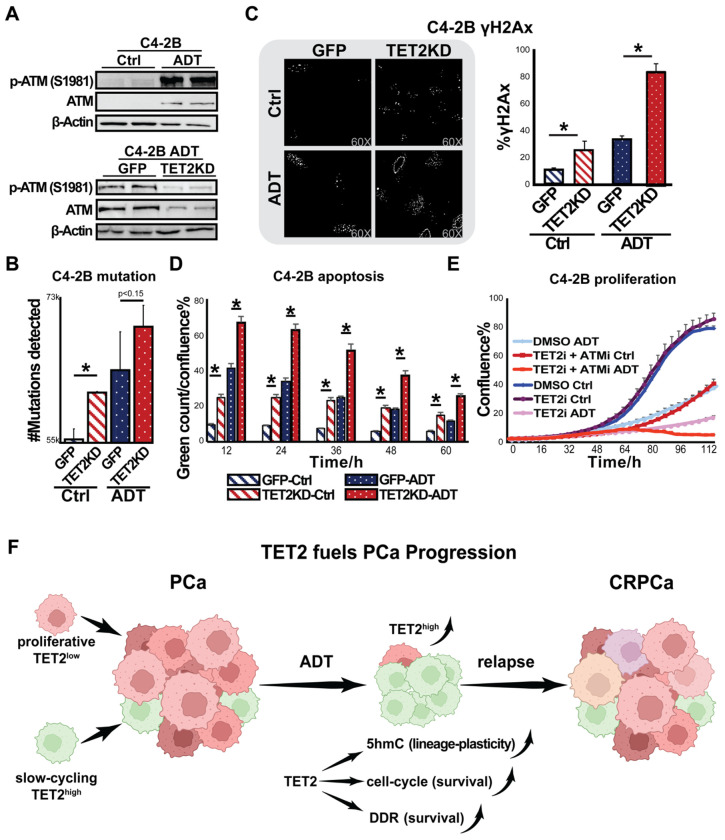
Advanced prostate cancer (PCa) frequently develops resistance to androgen deprivation therapy through various mechanisms including lineage plasticity. Slow-cycling cells (SCCs) have emerged as key players in adaptive responses to therapy, yet their role in PCa remains unclear. Through in silico analysis of single-cell RNA sequencing (scRNA-seq) data, we discovered that SCCs are enriched during pivotal stages of PCa progression, including the transition from androgen-dependent to castration-resistant states and the emergence of neuroendocrine PCa (NEPC). Using a tetracycline-inducible H2BeGFP reporter system, we confirmed SCC enrichment following androgen deprivation in both in vitro and in vivo models. Furthermore, we identified TET2 as a key regulator of SCCs, with its expression upregulated by androgen deprivation and positively correlated with SCC signature scores in PCa. Genome-wide 5-hydroxymethylcytosine (5hmC) profiling revealed increased hydroxymethylation after androgen deprivation, while TET2 knockdown reduced 5hmC levels at specific loci. Functional studies demonstrated that TET2 governs SCC maintenance, cell cycle progression, and DNA damage repair. Targeting TET2, either alone or in combination with an ATM inhibitor, significantly suppressed tumor growth, highlighting TET2 as a promising therapeutic target. Our study provides the first single-nucleotide resolution map of 5hmC dynamics in PCa, identifies a cell state driving epigenetic rewiring, and underscores the transformative potential of novel therapeutic strategies for advanced PCa.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous