

This is a preprint.
ATG2A engages Rab1a and ARFGAP1 positive membranes during autophagosome biogenesis
- PMID: 40196537
- PMCID: PMC11974814
- DOI: 10.1101/2025.03.24.645038
ATG2A engages Rab1a and ARFGAP1 positive membranes during autophagosome biogenesis
Abstract
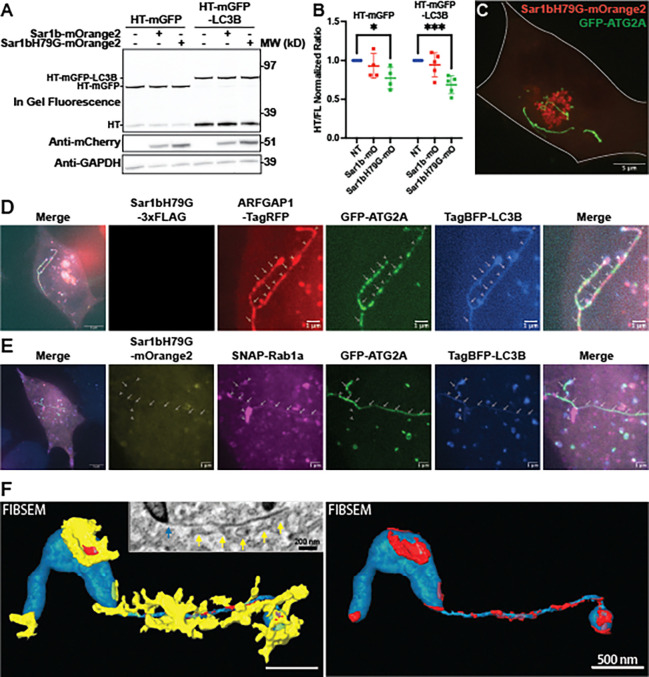
Autophagosomes form from seed membranes that expand through bulk-lipid transport via the bridge-like lipid transporter ATG2. The origins of the seed membranes and their relationship to the lipid transport machinery are poorly understood. Using proximity labeling and a variety of fluorescence microscopy techniques, we show that ATG2A localizes to extra-Golgi ARFGAP1 puncta during autophagosome biogenesis. ARFGAP1 itself is dispensable during macroautophagy, but among other proteins associating to these membranes, we find that Rab1 is essential. ATG2A co-immunoprecipitates strongly with Rab1a, and siRNA-mediated depletion of Rab1 blocks autophagy downstream of LC3B lipidation, similar to ATG2A depletion. Further, when either autophagosome formation or the early secretory pathway is perturbed, ARFGAP1 and Rab1a accumulate at ectopic locations with autophagic machinery. Our results suggest that ATG2A engages a Rab1a complex on select early secretory membranes at an early stage in autophagosome biogenesis.
Conflict of interest statement
Competing Interest Statement: FS and JB filed patent applications with the U.S. patent office covering FLASH-PAINT. JB has licensed IP to Bruker Corp. and Hamamatsu Photonics. JB is a founder of panluminate, Inc.
Figures



References
-
- Osawa T. et al. Atg2 mediates direct lipid transfer between membranes for autophagosome formation. Nat Struct Mol Biol 26, 281–288 (2019). - PubMed
-
- Osawa T., Ishii Y. & Noda N.N. Human ATG2B possesses a lipid transfer activity which is accelerated by negatively charged lipids and WIPI4. Genes Cells 25, 65–70 (2020). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources