

Dynamics of Spatial Organization of Bacterial Communities in a Tunable Flow Gut Microbiome-on-a-Chip
- PMID: 40201941
- PMCID: PMC12087827
- DOI: 10.1002/smll.202410258
Dynamics of Spatial Organization of Bacterial Communities in a Tunable Flow Gut Microbiome-on-a-Chip
Abstract
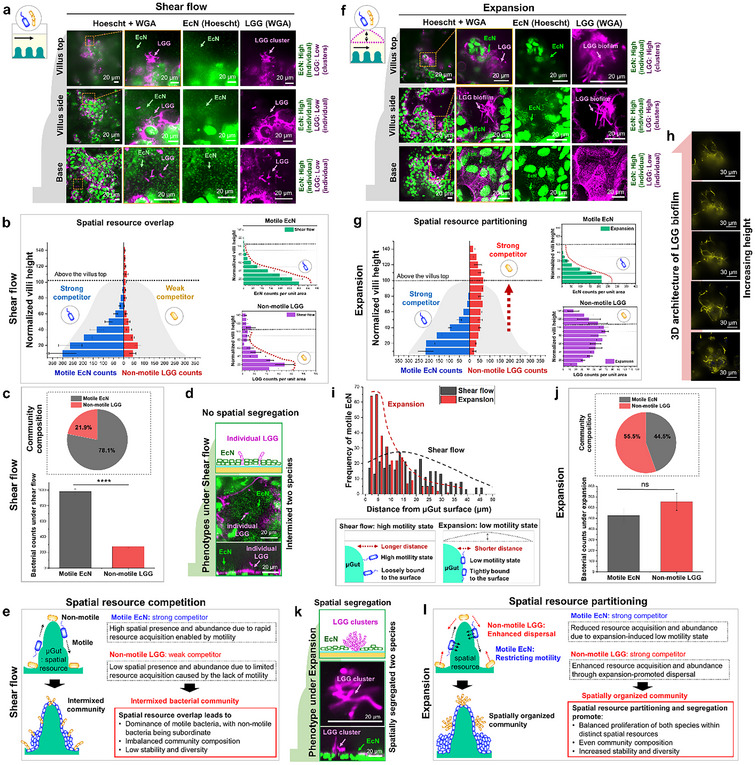
The human intestine, a biomechanically active organ, generates cyclic mechanical forces crucial for maintaining its health and functions. Yet, the physiological impact of these forces on gut microbiota dynamics remains largely unexplored. In this study, we investigate how cyclic intestinal motility influences the dynamics of gut microbial communities within a 3D gut-like structure (µGut). To enable the study, a tunable flow Gut Microbiome-on-a-Chip (tfGMoC) is developed that recapitulates the cyclic expansion and compression of intestinal motility while allowing high-magnification imaging of microbial communities within a 3D stratified, biomimetic gut epithelium. Using deep learning-based microbial analysis, it is found that hydrodynamic forces organize microbial communities by promoting distinct spatial exploration behaviors in microorganisms with varying motility characteristics. Empirical evidence demonstrates the impact of gut motility forces in maintaining a balanced gut microbial composition, enhancing both the diversity and stability of the community - key factors for a healthy microbiome. This study, leveraging the new tfGMoC platform, uncovers previously unknown effects of intestinal motility on modulating gut microbial behaviors and community organizations. This will be critical for a deeper understanding of host-microbiome interactions in the emerging field of microbiome therapeutics.
© 2025 The Author(s). Small published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Dissecting Gut-Microbial Community Interactions using a Gut Microbiome-on-a-Chip.Adv Sci (Weinh). 2024 May;11(20):e2302113. doi: 10.1002/advs.202302113. Epub 2024 Feb 27. Adv Sci (Weinh). 2024. PMID: 38414327 Free PMC article.
-
Genetic hypogonadal mouse model reveals niche-specific influence of reproductive axis and sex on intestinal microbial communities.Biol Sex Differ. 2023 Nov 6;14(1):79. doi: 10.1186/s13293-023-00564-1. Biol Sex Differ. 2023. PMID: 37932822 Free PMC article.
-
A complex human gut microbiome cultured in an anaerobic intestine-on-a-chip.Nat Biomed Eng. 2019 Jul;3(7):520-531. doi: 10.1038/s41551-019-0397-0. Epub 2019 May 13. Nat Biomed Eng. 2019. PMID: 31086325 Free PMC article.
-
Single-cell analysis and spatial resolution of the gut microbiome.Front Cell Infect Microbiol. 2023 Oct 4;13:1271092. doi: 10.3389/fcimb.2023.1271092. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37860069 Free PMC article. Review.
-
Gut-on-a-Chip for the Analysis of Bacteria-Bacteria Interactions in Gut Microbial Community: What Would Be Needed for Bacterial Co-Culture Study to Explore the Diet-Microbiota Relationship?Nutrients. 2023 Feb 23;15(5):1131. doi: 10.3390/nu15051131. Nutrients. 2023. PMID: 36904133 Free PMC article. Review.
References
-
- Consortium H. M. P., Nature 2012, 486, 207. - PubMed
-
- a) Kashyap P. C., Marcobal A., Ursell L. K., Larauche M., Duboc H., Earle K. A., Sonnenburg E. D., Ferreyra J. A., Higginbottom S. K., Million M., Tache Y., Pasricha P. J., Knight R., Farrugia G., Sonnenburg J. L., Gastroenterology 2013, 144, 967; - PMC - PubMed
- b) Touw K., Ringus D. L., Hubert N., Wang Y., Leone V. A., Nadimpalli A., Theriault B. R., Huang Y. E., Tune J. D., Herring P. B., Farrugia G., Kashyap P. C., Antonopoulos D. A., Chang E. B., Physiol. Rep. 2017, 5, 13182; - PMC - PubMed
- c) Murakami T., Kamada K., Mizushima K., Higashimura Y., Katada K., Uchiyama K., Handa O., Takagi T., Naito Y., Itoh Y., Digestion 2017, 95, 55; - PubMed
- d) Kim H. J., Li H., Collins J. J., Ingber D. E., Proc. Natl. Acad. Sci. USA 2016, 113, E7. - PMC - PubMed
-
- a) Persat A., Nadell C. D., Kim M. K., Ingremeau F., Siryaporn A., Drescher K., Wingreen N. S., Bassler B. L., Gitai Z., Stone H. A., Cell 2015, 161, 988; - PMC - PubMed
- b) Dufrêne Y. F., Persat A., Nat. Rev. Microbiol. 2020, 18, 227; - PubMed
- c) Rusconi R., Garren M., Stocker R., Annu. Rev. Biophys. 2014, 43, 65; - PMC - PubMed
- d) Cremer J., Segota I., Yang C. Y., Arnoldini M., Sauls J. T., Zhang Z., Gutierrez E., Groisman A., Hwa T., Proc. Natl. Acad. Sci. USA 2016, 113, 11414. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
