

Small molecules restore mutant mitochondrial DNA polymerase activity
- PMID: 40205042
- PMCID: PMC12158775
- DOI: 10.1038/s41586-025-08856-9
Small molecules restore mutant mitochondrial DNA polymerase activity
Abstract
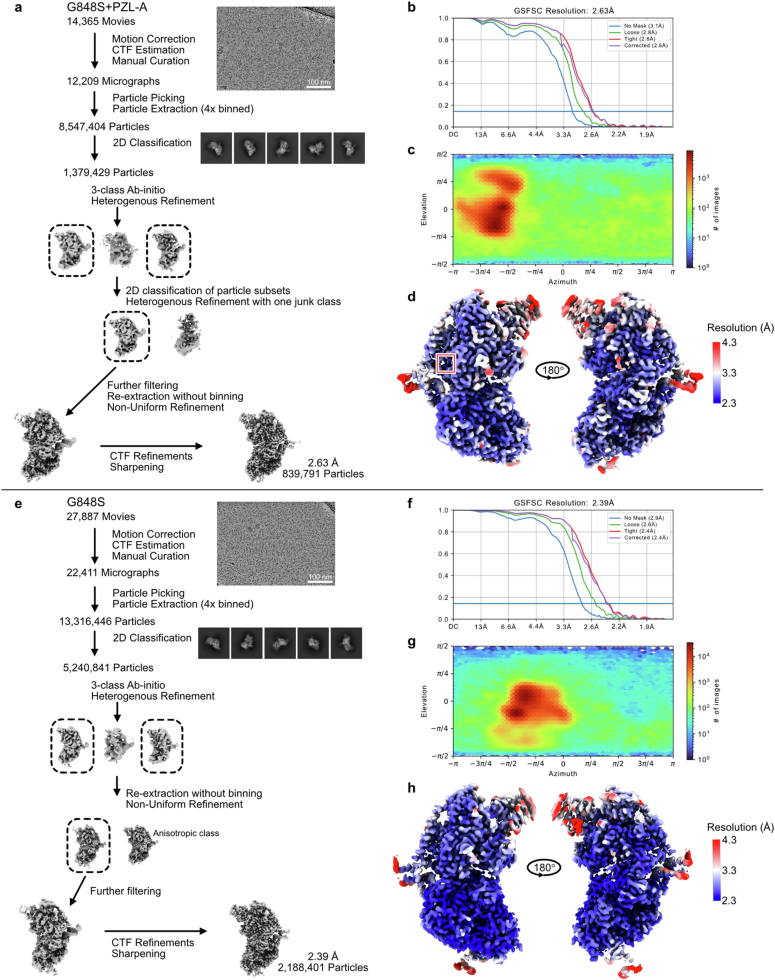
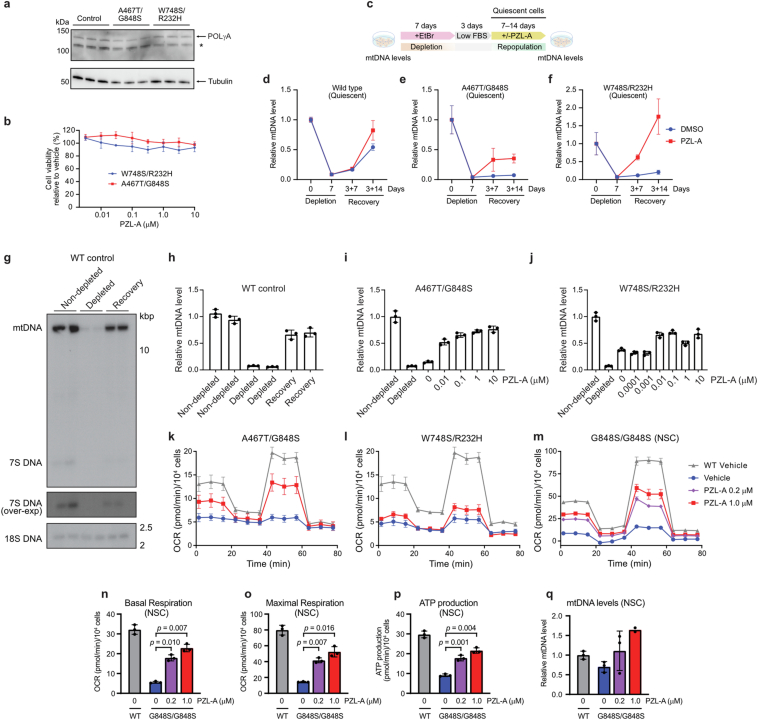
Mammalian mitochondrial DNA (mtDNA) is replicated by DNA polymerase γ (POLγ), a heterotrimeric complex consisting of a catalytic POLγA subunit and two accessory POLγB subunits1. More than 300 mutations in POLG, the gene encoding the catalytic subunit, have been linked to severe, progressive conditions with high rates of morbidity and mortality, for which no treatment exists2. Here we report on the discovery and characterization of PZL-A, a first-in-class small-molecule activator of mtDNA synthesis that is capable of restoring function to the most common mutant variants of POLγ. PZL-A binds to an allosteric site at the interface between the catalytic POLγA subunit and the proximal POLγB subunit, a region that is unaffected by nearly all disease-causing mutations. The compound restores wild-type-like activity to mutant forms of POLγ in vitro and activates mtDNA synthesis in cells from paediatric patients with lethal POLG disease, thereby enhancing biogenesis of the oxidative phosphorylation machinery and cellular respiration. Our work demonstrates that a small molecule can restore function to mutant DNA polymerases, offering a promising avenue for treating POLG disorders and other severe conditions linked to depletion of mtDNA.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: M.F., C.M.G., G.M.B. and N.-G.L. are co-founders of Pretzel Therapeutics. M.F., C.M.G., B.M., X.Z. and N.-G.L. are shareholders of Pretzel Therapeutics and have received consulting fees. S.V. has received consulting fees from Pretzel Therapeutics. M.S., C.G., A.V.G., J.M.F., Y.S., S.J.K., L.A., G.M.B., S.E., C.P.-H., T.A.K., B.K.-M., C.P., J.E.J., X.X. and S.G. are full-time employees of Pretzel Therapeutics and may hold stock or stock options as part of their compensation. V.P. is a former employee of Pretzel Therapeutics. P.S.C., A.M.G., G.K. and J.G. are consultants for Pretzel Therapeutics and hold stock or stock options as part of their compensation. The other authors declare no competing interests.
Figures











References
-
- Falkenberg, M., Larsson, N. G. & Gustafsson, C. M. Replication and transcription of human mitochondrial DNA. Annu. Rev. Biochem.93, 47–77 (2024). - PubMed
-
- Suomalainen, A. & Nunnari, J. Mitochondria at the crossroads of health and disease. Cell187, 2601–2627 (2024). - PubMed
-
- Gorman, G. S. et al. Mitochondrial diseases. Nat. Rev. Dis. Primers2, 16080 (2016). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
