

Complete sequencing of ape genomes
- PMID: 40205052
- PMCID: PMC12058530
- DOI: 10.1038/s41586-025-08816-3
Complete sequencing of ape genomes
Abstract
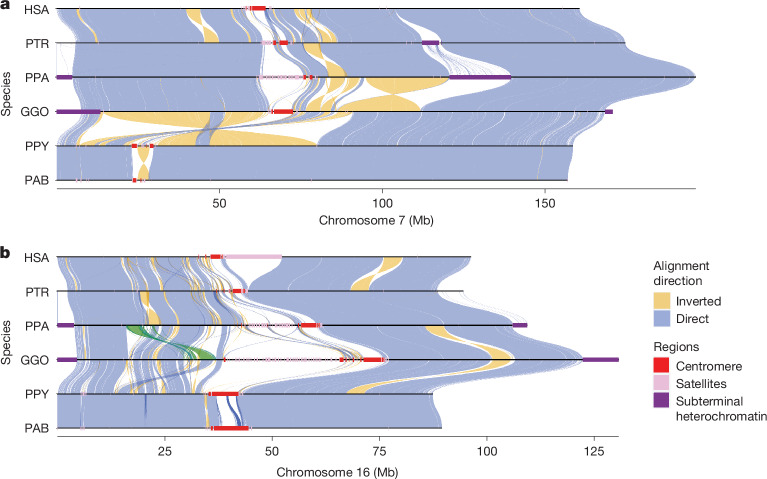
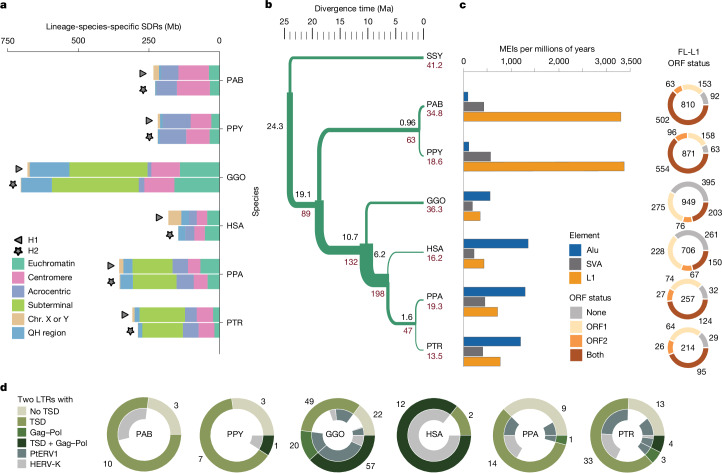
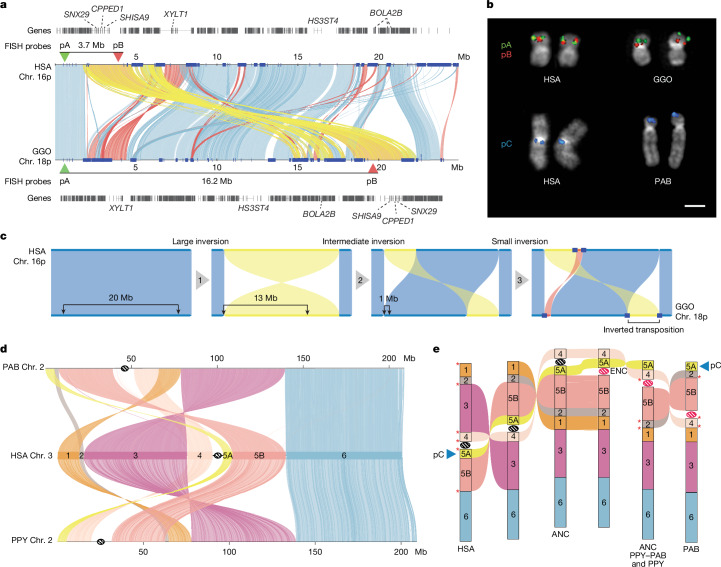
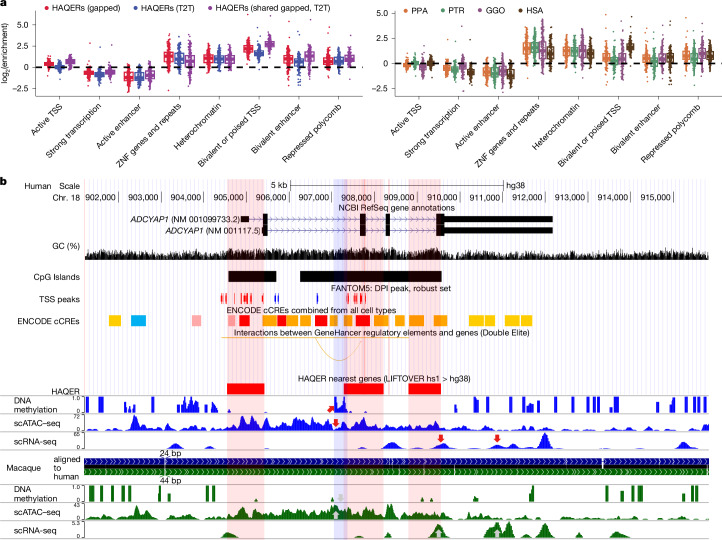
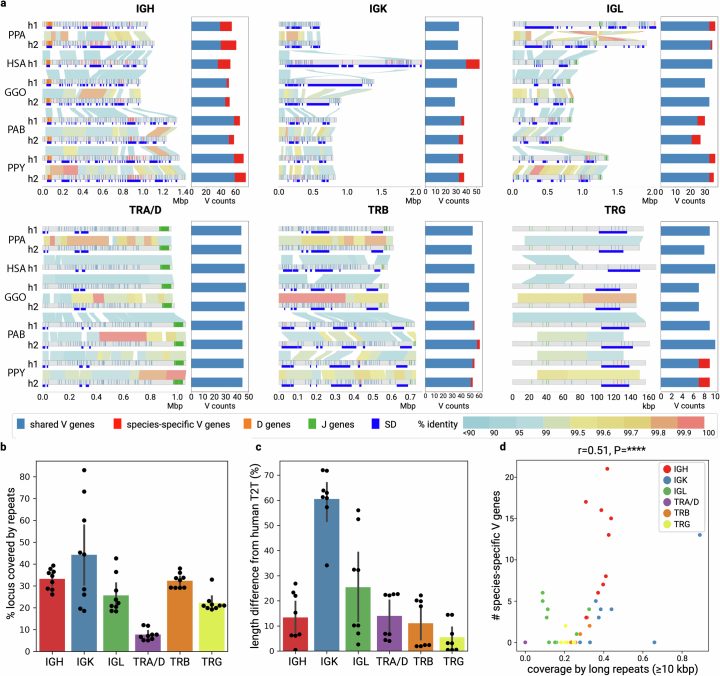
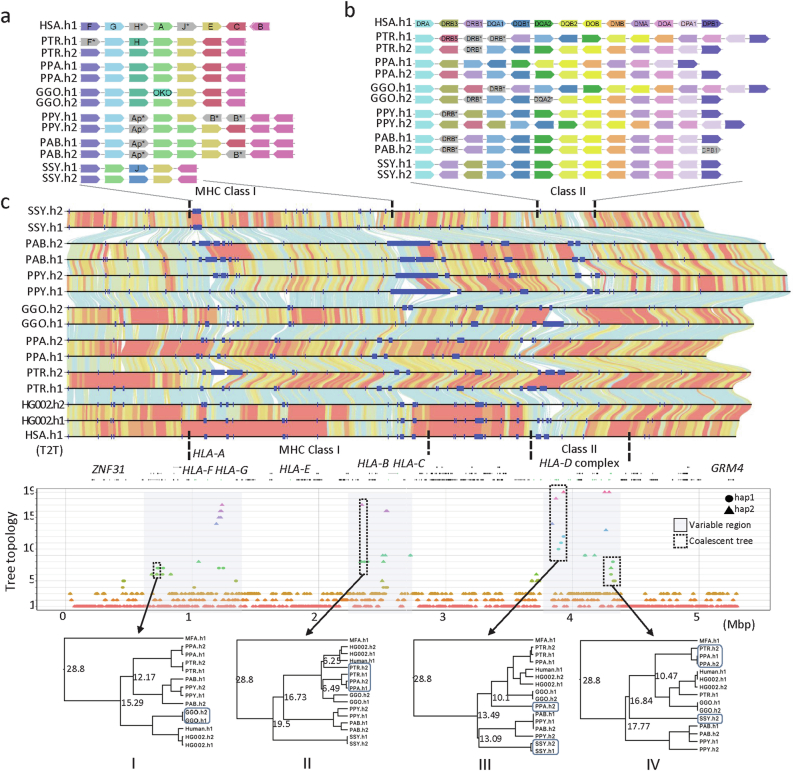
The most dynamic and repetitive regions of great ape genomes have traditionally been excluded from comparative studies1-3. Consequently, our understanding of the evolution of our species is incomplete. Here we present haplotype-resolved reference genomes and comparative analyses of six ape species: chimpanzee, bonobo, gorilla, Bornean orangutan, Sumatran orangutan and siamang. We achieve chromosome-level contiguity with substantial sequence accuracy (<1 error in 2.7 megabases) and completely sequence 215 gapless chromosomes telomere-to-telomere. We resolve challenging regions, such as the major histocompatibility complex and immunoglobulin loci, to provide in-depth evolutionary insights. Comparative analyses enabled investigations of the evolution and diversity of regions previously uncharacterized or incompletely studied without bias from mapping to the human reference genome. Such regions include newly minted gene families in lineage-specific segmental duplications, centromeric DNA, acrocentric chromosomes and subterminal heterochromatin. This resource serves as a comprehensive baseline for future evolutionary studies of humans and our closest living ape relatives.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: E.E.E. is a scientific advisory board member of Variant Bio. C.T.W. is a co-founder and Chief Scientific Officer of Clareo Biosciences. W.L. is a co-founder and Chief Technology Officer of Clareo Biosciences. The other authors declare no competing interests.
Figures

Update of
-
Complete sequencing of ape genomes.bioRxiv [Preprint]. 2024 Oct 5:2024.07.31.605654. doi: 10.1101/2024.07.31.605654. bioRxiv. 2024. Update in: Nature. 2025 May;641(8062):401-418. doi: 10.1038/s41586-025-08816-3. PMID: 39131277 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 HG012344/HG/NHGRI NIH HHS/United States
- R01 MH120295/MH/NIMH NIH HHS/United States
- UH3 AG064706/AG/NIA NIH HHS/United States
- R35 GM146886/GM/NIGMS NIH HHS/United States
- R01 HG010329/HG/NHGRI NIH HHS/United States
- R01 AG056169/AG/NIA NIH HHS/United States
- R01 GM123312/GM/NIGMS NIH HHS/United States
- T15 LM007093/LM/NLM NIH HHS/United States
- U24 HG007497/HG/NHGRI NIH HHS/United States
- R01 HG002385/HG/NHGRI NIH HHS/United States
- P20 GM139769/GM/NIGMS NIH HHS/United States
- U24 HG010263/HG/NHGRI NIH HHS/United States
- R35 GM133747/GM/NIGMS NIH HHS/United States
- P51 OD011092/OD/NIH HHS/United States
- R56 AG056169/AG/NIA NIH HHS/United States
- P01 AI152999/AI/NIAID NIH HHS/United States
- R35 HG011332/HG/NHGRI NIH HHS/United States
- R35 GM133600/GM/NIGMS NIH HHS/United States
- U19 AG023122/AG/NIA NIH HHS/United States
- R01 HG007352/HG/NHGRI NIH HHS/United States
- R35 GM142916/GM/NIGMS NIH HHS/United States
- R01 HG010169/HG/NHGRI NIH HHS/United States
- U41 HG007234/HG/NHGRI NIH HHS/United States
- R35 GM146926/GM/NIGMS NIH HHS/United States
- R35 GM151348/GM/NIGMS NIH HHS/United States
- R35 GM151945/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
