

Most L1CAM Is not Associated with Extracellular Vesicles in Human Biofluids and iPSC-Derived Neurons
- PMID: 40210837
- PMCID: PMC12289729
- DOI: 10.1007/s12035-025-04909-2
Most L1CAM Is not Associated with Extracellular Vesicles in Human Biofluids and iPSC-Derived Neurons
Abstract
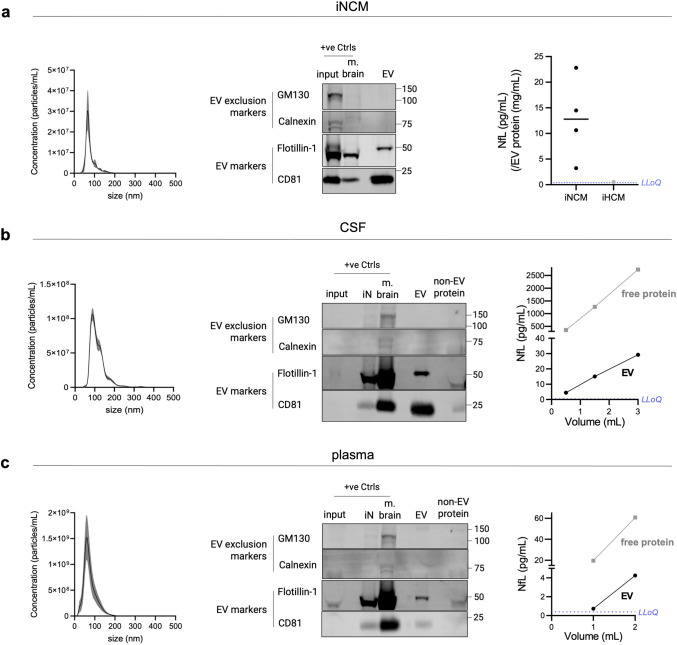
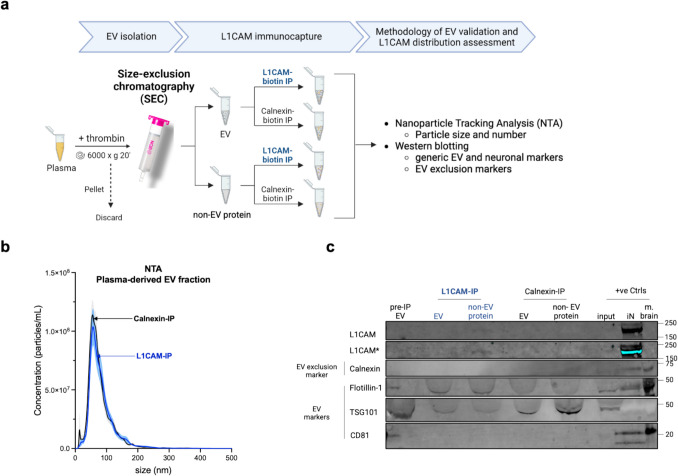
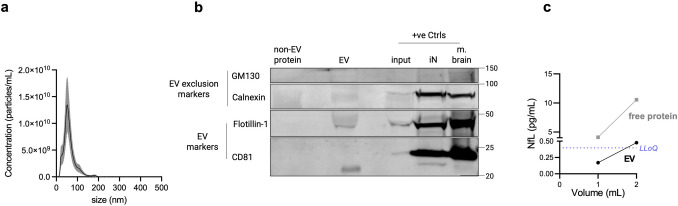
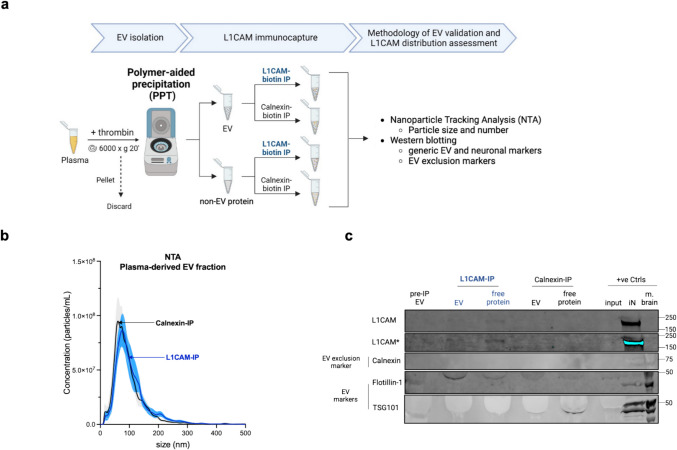
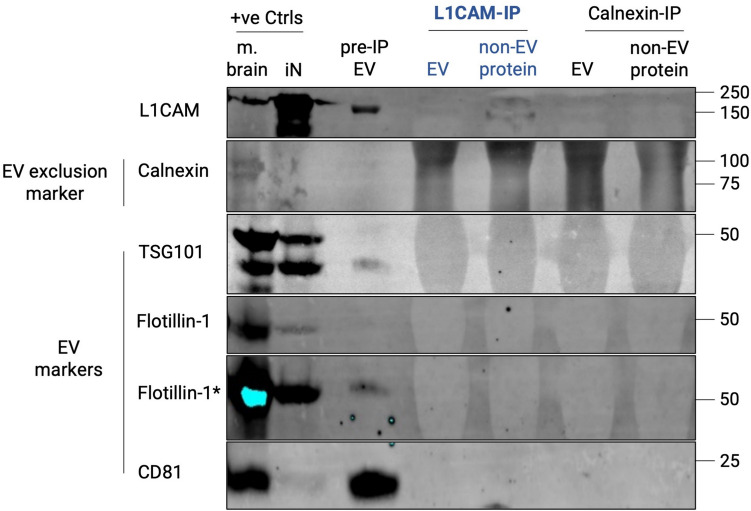
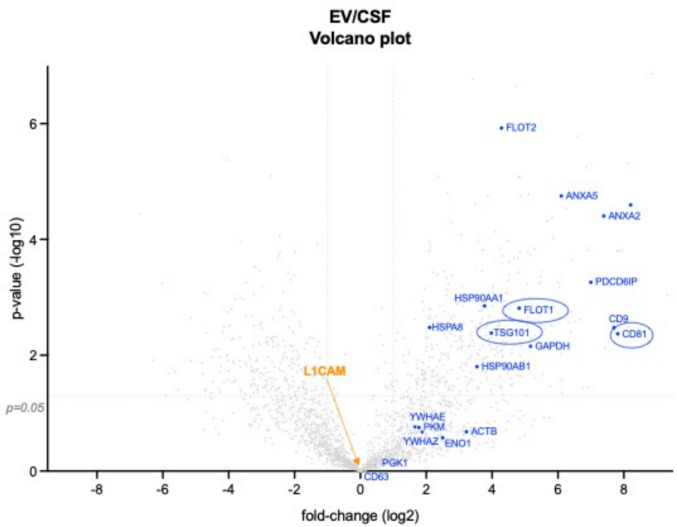
Transmembrane L1 cell adhesion molecule (L1CAM) is widely used as a marker to enrich for neuron-derived extracellular vesicles (EVs), especially in plasma. However, this approach lacks sufficient robust validation. This study aimed to assess whether human biofluids are indeed enriched for EVs, particularly neuron-derived EVs, by L1CAM immunoaffinity, utilizing multiple sources (plasma, CSF, conditioned media from iPSC-derived neurons [iNCM]) and different methods (mass spectrometry [MS], nanoparticle tracking analysis [NTA]). Following a systematic multi-step validation approach, we confirmed isolation of generic EV populations using size-exclusion chromatography (SEC) and polymer-aided precipitation (PPT)-two most commonly applied EV isolation methods-from all sources. Neurofilament light (NfL) was detected in both CSF and blood-derived EVs, indicating their neuronal origin. However, L1CAM immunoprecipitation did not yield enrichment of L1CAM in EV fractions. Instead, it was predominantly found in its free-floating form. Additionally, MS-based proteomic analysis of CSF-derived EVs also did not show L1CAM enrichment. Our study validates EV isolation from diverse biofluid sources by several isolation approaches and confirms that some EV subpopulations in human biofluids are of neuronal origin. Thorough testing across multiple sources by different orthogonal methods, however, does not support L1CAM as a marker to reliably enrich for a specific subpopulation of EVs, particularly of neuronal origin.
Keywords: Biomarkers; Blood; Cerebrospinal fluid; Extracellular vesicles; Immunoprecipitation; Isolation methods; L1CAM; Neuron.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Informed Consent Statement: Informed consent was obtained from all subjects involved this study wherever necessary. Ethics Approval: This study was performed in line with the principles of the Declaration of Helsinki and approved by the Ethics Committee of the University of Tuebingen (199/2011BO1). Conflict of Interest: The authors declare no competing interests.
Figures
Similar articles
-
In-Depth Characterization of L1CAM+ Extracellular Vesicles as Potential Biomarkers for Anti-CD20 Therapy Response in Relapsing-Remitting Multiple Sclerosis.Int J Mol Sci. 2025 Jul 25;26(15):7213. doi: 10.3390/ijms26157213. Int J Mol Sci. 2025. PMID: 40806340 Free PMC article.
-
Examination of the enrichment of neuronal extracellular vesicles from cell conditioned media and human plasma using an anti-NCAM immunocapture bead approach.bioRxiv [Preprint]. 2025 May 14:2025.05.13.653678. doi: 10.1101/2025.05.13.653678. bioRxiv. 2025. Update in: BMC Methods. 2025;2(1):12. doi: 10.1186/s44330-025-00034-7. PMID: 40463084 Free PMC article. Updated. Preprint.
-
Examination of the enrichment of neuronal extracellular vesicles from cell conditioned media and human plasma using an anti-NCAM immunocapture bead approach.BMC Methods. 2025;2(1):12. doi: 10.1186/s44330-025-00034-7. Epub 2025 Jul 1. BMC Methods. 2025. PMID: 40606859 Free PMC article.
-
Central nervous system-derived extracellular vesicles: the next generation of neural circulating biomarkers?Transl Neurodegener. 2024 Jun 19;13(1):32. doi: 10.1186/s40035-024-00418-9. Transl Neurodegener. 2024. PMID: 38898538 Free PMC article. Review.
-
Extracellular Vesicles for the Diagnosis of Parkinson's Disease: Systematic Review and Meta-Analysis.Mov Disord. 2023 Sep;38(9):1585-1597. doi: 10.1002/mds.29497. Epub 2023 Jul 14. Mov Disord. 2023. PMID: 37449706
Cited by
-
Neuron-derived extracellular vesicles in plasma present a potential non-invasive biomarker for Huntingtin protein and RNA assessment in Huntington disease.bioRxiv [Preprint]. 2025 Jul 21:2025.07.17.665403. doi: 10.1101/2025.07.17.665403. bioRxiv. 2025. PMID: 40777434 Free PMC article. Preprint.
-
Differential microRNA expression profiling of peripheral blood L1CAM neural-enriched and bulk extracellular vesicles in individuals with bipolar disorder.medRxiv [Preprint]. 2025 Jun 6:2025.06.06.25329080. doi: 10.1101/2025.06.06.25329080. medRxiv. 2025. PMID: 40502571 Free PMC article. Preprint.
-
Serum sEV miRNAs as Biomarkers in Myelin Oligodendrocyte Glycoprotein Antibody-Associated Disease.Mol Neurobiol. 2025 Sep;62(9):12280-12295. doi: 10.1007/s12035-025-04932-3. Epub 2025 May 19. Mol Neurobiol. 2025. PMID: 40388105 Free PMC article.
References
-
- Cocozza F, Grisard E, Martin-Jaular L, Mathieu M (2020) Théry C. SnapShot: extracellular vesicles. Cell 182(1):262 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
