

The Spatial Transcriptional Activity of Hepatic TCF7L2 Regulates Zonated Metabolic Pathways that Contribute to Liver Fibrosis
- PMID: 40210847
- PMCID: PMC11986045
- DOI: 10.1038/s41467-025-58714-5
The Spatial Transcriptional Activity of Hepatic TCF7L2 Regulates Zonated Metabolic Pathways that Contribute to Liver Fibrosis
Abstract
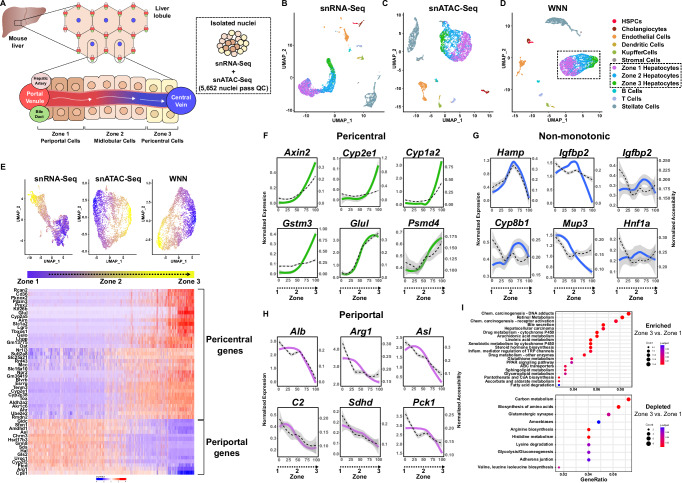
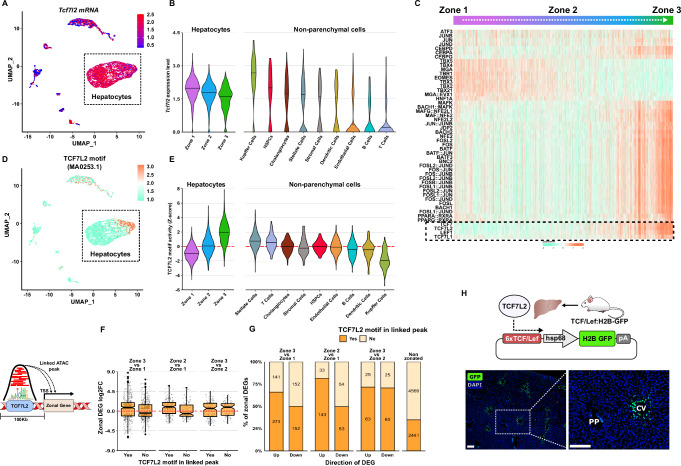
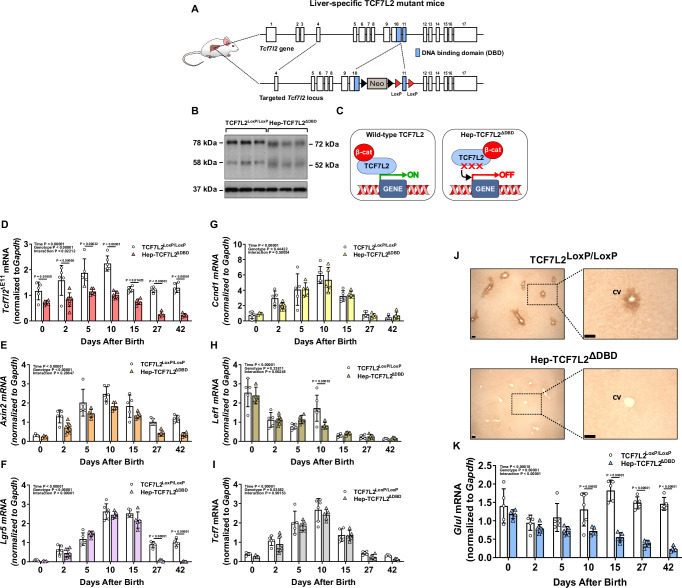
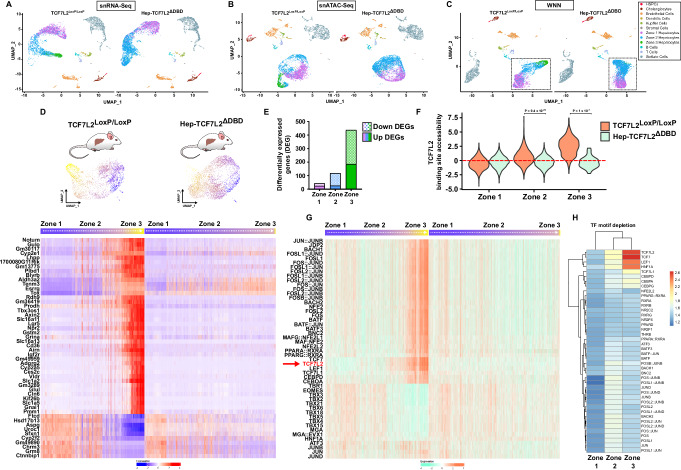
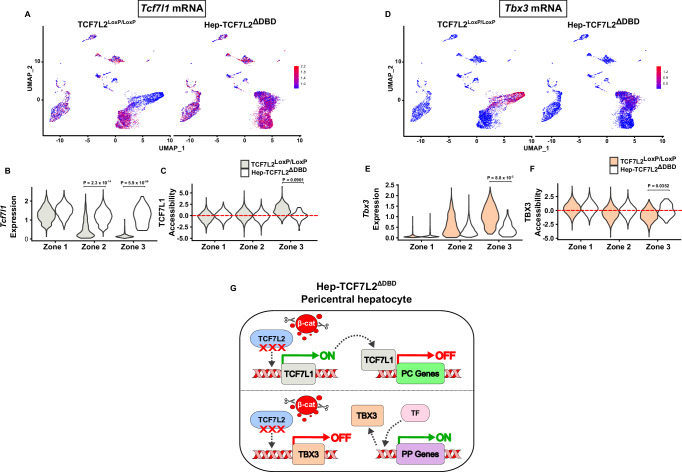
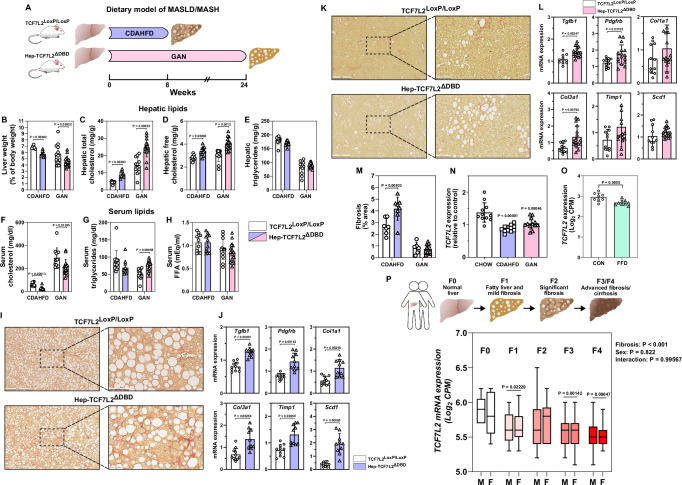
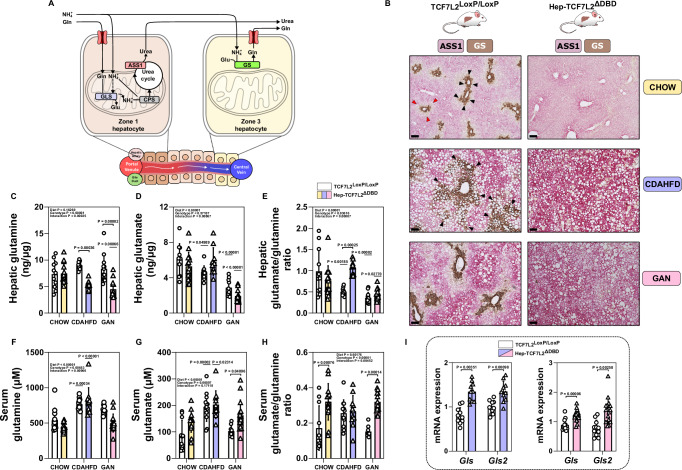
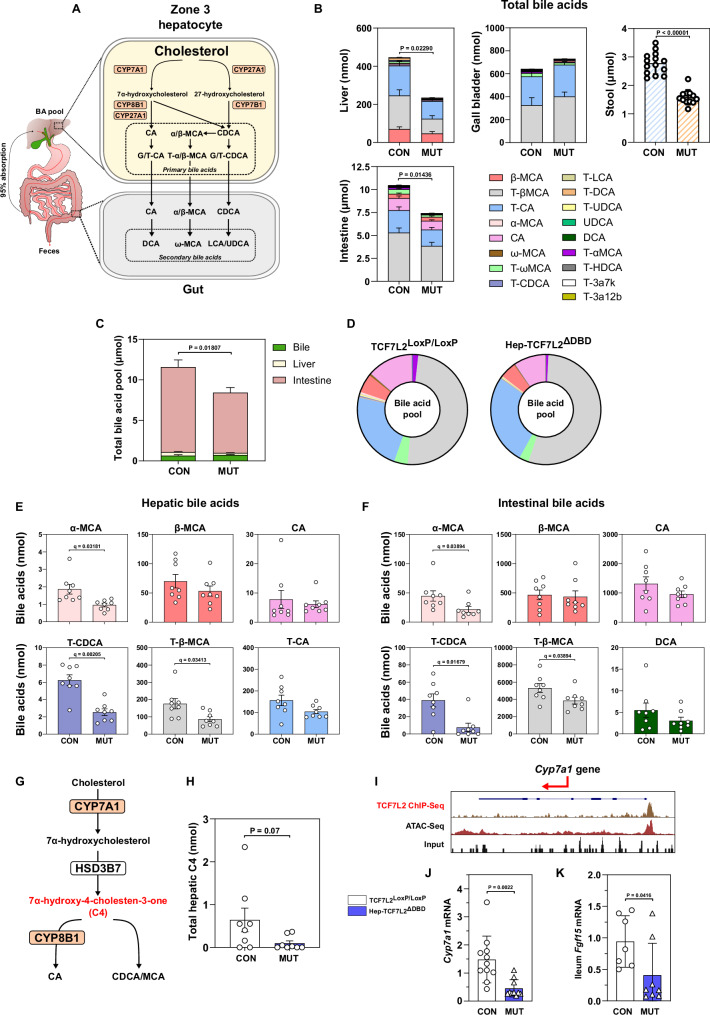
The molecular mechanisms regulating the zonal distribution of metabolism in liver are incompletely understood. Here we use single nuclei genomics techniques to examine the spatial transcriptional function of transcription factor 7-like 2 (TCF7L2) in mouse liver, and determine the consequences of TCF7L2 transcriptional inactivation on the metabolic architecture of the liver and the function of zonated metabolic pathways. We report that while Tcf7l2 mRNA expression is ubiquitous across the liver lobule, accessibility of the consensus TCF/LEF DNA binding motif is restricted to pericentral (PC) hepatocytes in zone 3. In mice expressing functionally inactive TCF7L2 in liver, PC hepatocyte-specific gene expression is absent, which we demonstrate promotes hepatic cholesterol accumulation, impaired bile acid synthesis, disruption to glutamine/glutamate homeostasis and pronounced dietary-induced hepatic fibrosis. In summary, TCF7L2 is a key regulator of hepatic zonal gene expression and regulates several zonated metabolic pathways that may contribute to the development of fibrotic liver disease.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
-
- Asrani, S. K. et al. Burden of liver diseases in the world. J. Hepatol.70, 151–171 (2019). - PubMed
-
- Rinella, M. E. et al. A multi-society Delphi consensus statement on new fatty liver disease nomenclature. J. Hepatol. 78, 1966–1986 2023. - PubMed
-
- Harrison, S. A. et al. Prospective evaluation of the prevalence of non-alcoholic fatty liver disease and steatohepatitis in a large middle-aged US cohort. J. Hepatol. 75, 284–291 2021. - PubMed
-
- Gebhardt, R. Metabolic zonation of the liver: regulation and implications for liver function. Pharm. Ther.53, 275–354 (1992). - PubMed
MeSH terms
Substances
Grants and funding
- R01 DK138340/DK/NIDDK NIH HHS/United States
- R01DK128247/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- T32 TR004544/TR/NCATS NIH HHS/United States
- TL1 TR002647/TR/NCATS NIH HHS/United States
- R01 DK128247/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
