

Epigenomic signatures of cis-regulatory elements in the developing mouse and pig forelimb
- PMID: 40210926
- PMCID: PMC11986027
- DOI: 10.1038/s41597-025-04946-5
Epigenomic signatures of cis-regulatory elements in the developing mouse and pig forelimb
Abstract
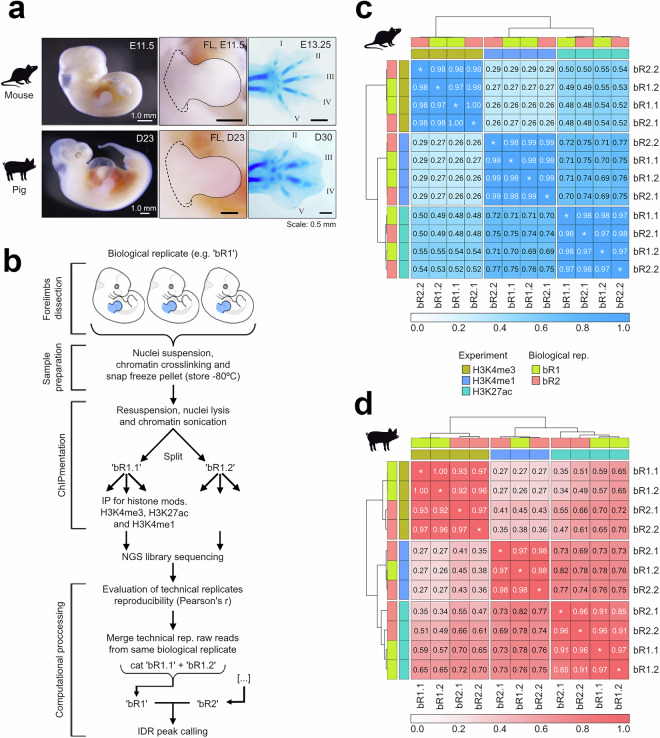
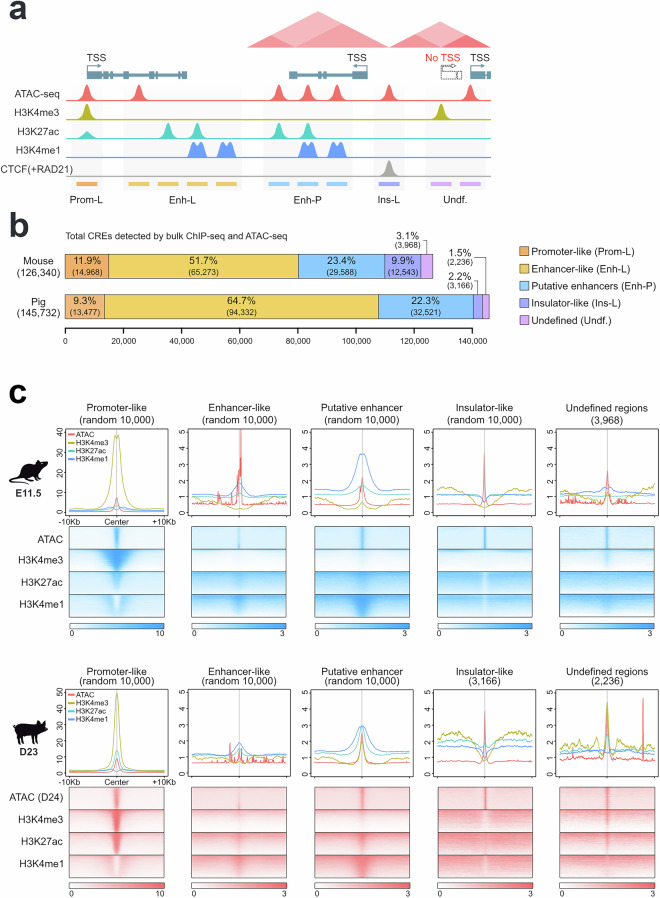
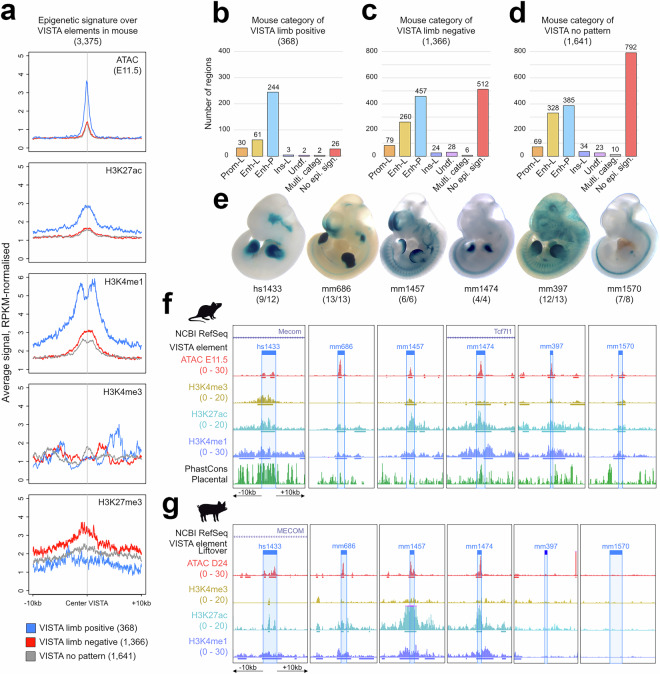
Cis-regulatory elements (CREs) orchestrate the spatiotemporal regulation of key transcriptional programs. These genomic regions-including promoters, enhancers, and insulators-play a crucial role during embryonic development, and their functional diversification is thought to contribute significantly to the morphological evolution of animal body plans. We performed chromatin immunoprecipitation for various histone modifications (H3K4me3, H3K27ac, and H3K4me1) from equivalent developmental stages of mouse (E11.5) and pig (day 23; D23) to identify active regulatory regions during forelimb development in both species. The overlap of these epigenomic signatures with the pattern of open chromatin allowed us to classify these putative regulatory regions into different chromatin states in mouse and pig limb primordia. Our profiling of the regulatory genome in mouse and pig limb buds offers a valuable resource in the evo-devo field for exploring mechanisms underlying the morphological evolution of the tetrapod limb.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
