

NETs activate the GAS6-AXL-NLRP3 axis in macrophages to drive morphine tolerance
- PMID: 40217343
- PMCID: PMC11992818
- DOI: 10.1186/s12964-025-02181-4
NETs activate the GAS6-AXL-NLRP3 axis in macrophages to drive morphine tolerance
Abstract
Background: The development of morphine tolerance presents a major clinical challenge in the effective management of severe pain. This study aims to explore the mechanisms underlying morphine tolerance from a novel perspective, with the ultimate goal of uncovering new insights and identifying promising therapeutic targets for its treatment.
Methods: C57BL/6J mice were used in the tail-flick test to evaluate morphine tolerance. Neutrophils derived from mouse bone marrow were employed to investigate the mechanisms underlying morphine-induced NETs formation. Bone marrow-derived macrophages (BMDMs) were harvested from the femur and tibia to study the role of NETs-induced inflammation in analgesic tolerance. Proinflammatory cytokines were measured using Western blotting and real-time PCR. The levels of NETs and the TLR7/9-NLRP3-related signaling pathway were assessed through Western blotting, real-time PCR, and ELISA. Confocal laser scanning microscopy was utilized to visualize NETs in the dorsal root ganglion (DRG) and in cells.
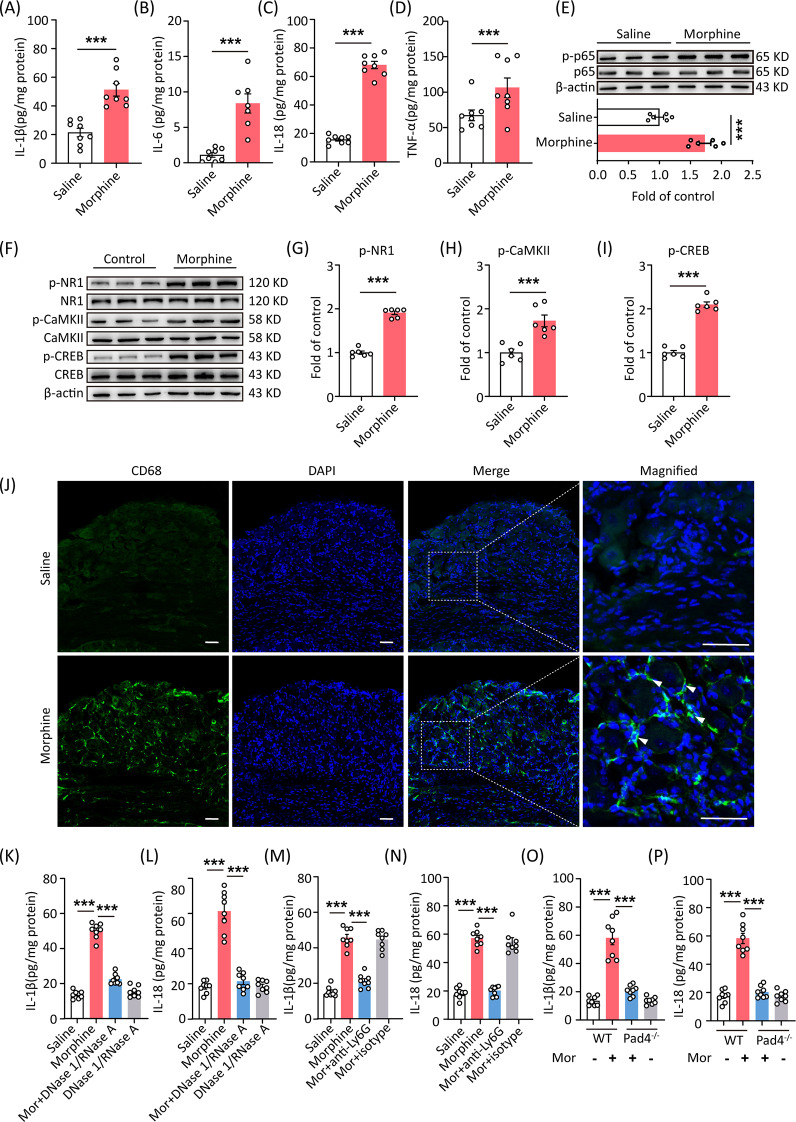
Results: Our experiments demonstrated that the levels of NETs in the plasma of patients using morphine for analgesia, as well as in morphine-tolerant animals, were significantly elevated. Genetic elimination of Pad4, neutrophil depletion, and treatment with DNase 1 and RNase A to disrupt NETs formation all effectively alleviated morphine tolerance. These findings indicate that NETs play a critical role in the development of morphine tolerance. Mechanistically, we discovered that morphine-induced NETs can be engulfed by macrophages through the GAS6-AXL axis, which subsequently triggers the activation of the TLR7/TLR9-mediated NLRP3 inflammasome, leading to significantly increased levels of IL-1β and IL-18, and ultimately contributing to tolerance. Deletion of Axl, Gas6, or Nlrp3 each significantly improved morphine tolerance. Furthermore, in the murine model, treatment with the IL-1 receptor antagonist anakinra and the IL-18 decoy receptor IL-18BP prevented the development of morphine tolerance.
Conclusions: This study identifies morphine-induced NETs as a key contributor to morphine tolerance, with the GAS6-AXL-TLR7/9 axis emerging as a potential therapeutic target. Strategies focused on disrupting NETs and modulating this axis may offer a promising approach to combat morphine tolerance.
Keywords: AXL; GAS6; Morphine tolerance; NETs; NLRP3.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The studies involving human participants were approved by the Ethics Committee of the First Affiliated Hospital of Nanjing Medical University (approval number: 2023-SR-031). All participants signed informed consent. All animal experiments were conducted according to protocols approved by the Animal Research Ethics Committee of Nanjing Medical University (approval number: IACUC-2203057-1). Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures






Similar articles
-
Gas6 induces AIM to suppress acute lung injury in mice by inhibiting NLRP3 inflammasome activation and inducing autophagy.Front Immunol. 2025 Feb 17;16:1523166. doi: 10.3389/fimmu.2025.1523166. eCollection 2025. Front Immunol. 2025. PMID: 40034700 Free PMC article.
-
Autophagy induced by AXL receptor tyrosine kinase alleviates acute liver injury via inhibition of NLRP3 inflammasome activation in mice.Autophagy. 2016 Dec;12(12):2326-2343. doi: 10.1080/15548627.2016.1235124. Autophagy. 2016. PMID: 27780404 Free PMC article.
-
Blocking ATP-sensitive potassium channel alleviates morphine tolerance by inhibiting HSP70-TLR4-NLRP3-mediated neuroinflammation.J Neuroinflammation. 2017 Nov 25;14(1):228. doi: 10.1186/s12974-017-0997-0. J Neuroinflammation. 2017. PMID: 29178967 Free PMC article.
-
Involvement of the NLRP3 inflammasome in the modulation of an LPS-induced inflammatory response during morphine tolerance.Drug Alcohol Depend. 2013 Sep 1;132(1-2):38-46. doi: 10.1016/j.drugalcdep.2012.12.022. Epub 2013 Jan 23. Drug Alcohol Depend. 2013. PMID: 23352192
-
A Functional Role of GAS6/TAM in Nonalcoholic Steatohepatitis Progression Implicates AXL as Therapeutic Target.Cell Mol Gastroenterol Hepatol. 2020;9(3):349-368. doi: 10.1016/j.jcmgh.2019.10.010. Epub 2019 Nov 2. Cell Mol Gastroenterol Hepatol. 2020. PMID: 31689560 Free PMC article.
References
MeSH terms
Substances
Grants and funding
- 81971047, 81773798, 82271252/the National Natural Science Foundation of China
- BK20191253/Natural Science Foundation of Jiangsu Province
- 2017NJMUCX004/Major Project of "Science and Technology Innovation Fund" of Nanjing Medical University
- BE2019732/Key R&D Program (Social Development) Project of Jiangsu Province
- YKK19170/Nanjing Special Fund for Health Science and Technology Development
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
