

A genome-wide screen identifies genes required for erythroid differentiation
- PMID: 40221460
- PMCID: PMC11993733
- DOI: 10.1038/s41467-025-58739-w
A genome-wide screen identifies genes required for erythroid differentiation
Abstract
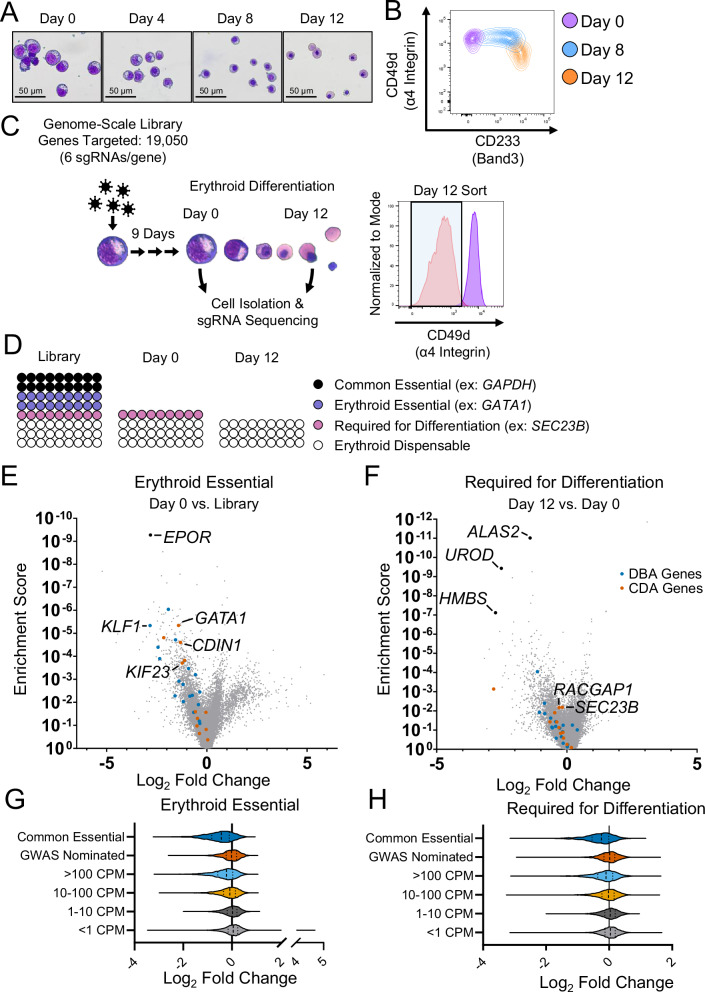
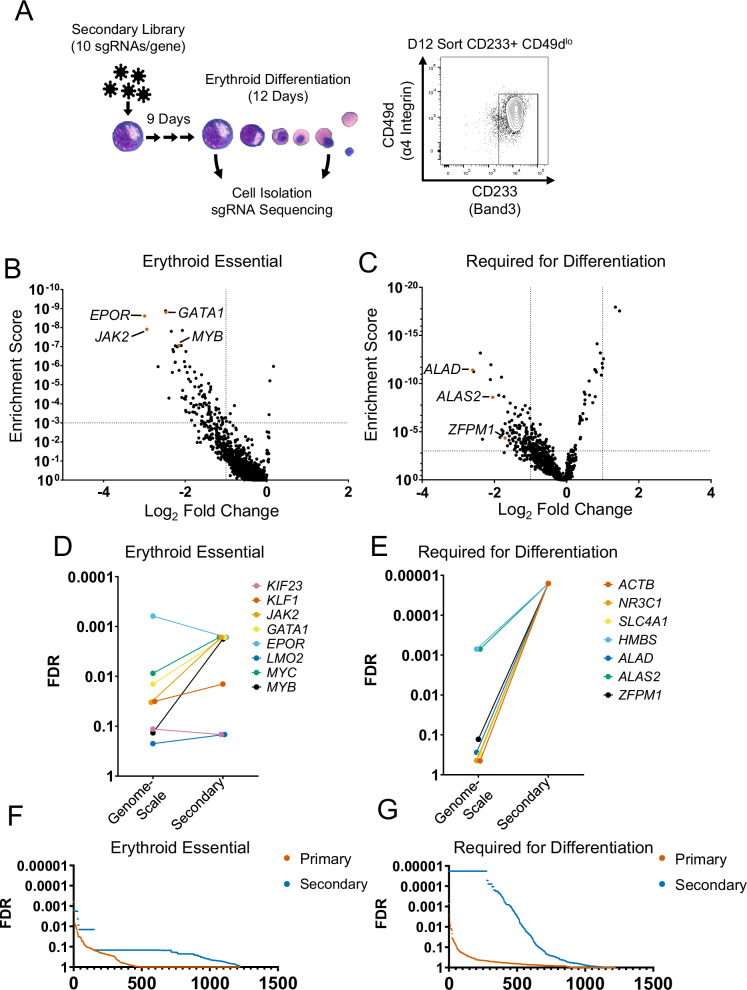
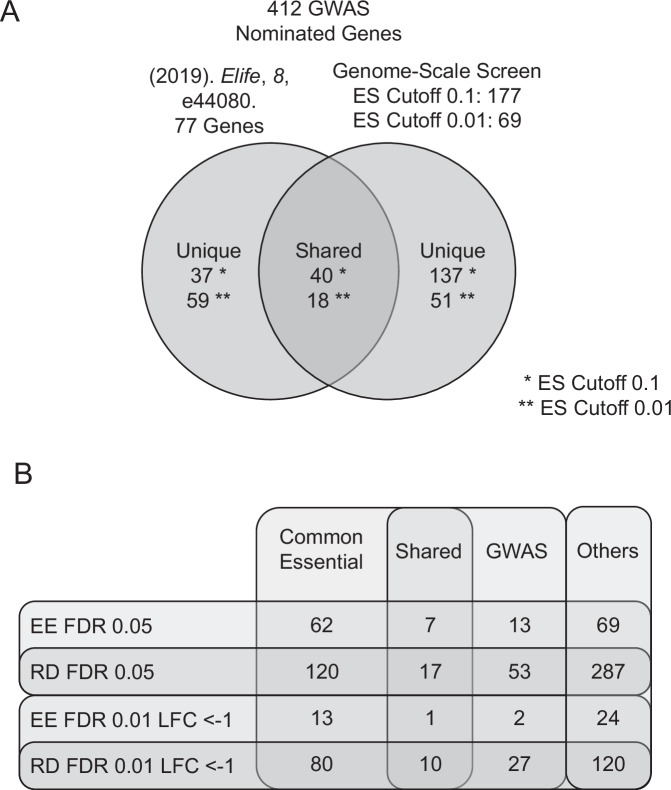
The complete array of genes required for terminal erythroid differentiation remains unknown. To address this knowledge gap, we perform a genome-scale CRISPR knock-out screen in the human erythroid progenitor cell line HUDEP-2 and validate candidate regulators of erythroid differentiation in a custom secondary screen. Comparison of sgRNA abundance in the CRISPR library, proerythroblasts, and orthochromatic erythroblasts, resulted in the identification of genes that are essential for proerythroblast survival and genes that are required for terminal erythroid differentiation. Among the top genes identified are known regulators of erythropoiesis, underscoring the validity of this screen. Notably, using a Log2 fold change of <-1 and false discovery rate of <0.01, the screen identified 277 genes that are required for terminal erythroid differentiation, including multiple genes not previously nominated through GWAS. NHLRC2, which was previously implicated in hemolytic anemia, was a highly ranked gene. We suggest that anemia due to NHLRC2 mutation results at least in part from a defect in erythroid differentiation. Another highly ranked gene in the screen is VAC14, which we validated for its requirement in erythropoiesis in vitro and in vivo. Thus, data from this CRISPR screen may help classify the underlying mechanisms that contribute to erythroid disorders.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: I.M. has received research funding from Genentech and Regeneron, and he is a member of Garuda Therapeutic’s scientific advisory board. Other authors declare no competing interests.
Figures



References
MeSH terms
Grants and funding
- R01 AI091627/AI/NIAID NIH HHS/United States
- U2C DK129445/DK/NIDDK NIH HHS/United States
- T32 GM007863/GM/NIGMS NIH HHS/United States
- F31 HL162544/HL/NHLBI NIH HHS/United States
- K08 DK127013/DK/NIDDK NIH HHS/United States
- P01 HL146372/HL/NHLBI NIH HHS/United States
- R01 HL157062/HL/NHLBI NIH HHS/United States
- U01 HL117658/HL/NHLBI NIH HHS/United States
- T32 GM007315/GM/NIGMS NIH HHS/United States
- TL1 DK136046/DK/NIDDK NIH HHS/United States
- T32-GM007315/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- P30 CA008748/CA/NCI NIH HHS/United States
- AI091627/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R01 HL148333/HL/NHLBI NIH HHS/United States
- U54 DK106829/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
