

Escherichia coli type I toxin TisB exclusively controls proton depolarization following antibiotic induced DNA damage
- PMID: 40229382
- PMCID: PMC11997105
- DOI: 10.1038/s41598-025-96136-x
Escherichia coli type I toxin TisB exclusively controls proton depolarization following antibiotic induced DNA damage
Abstract
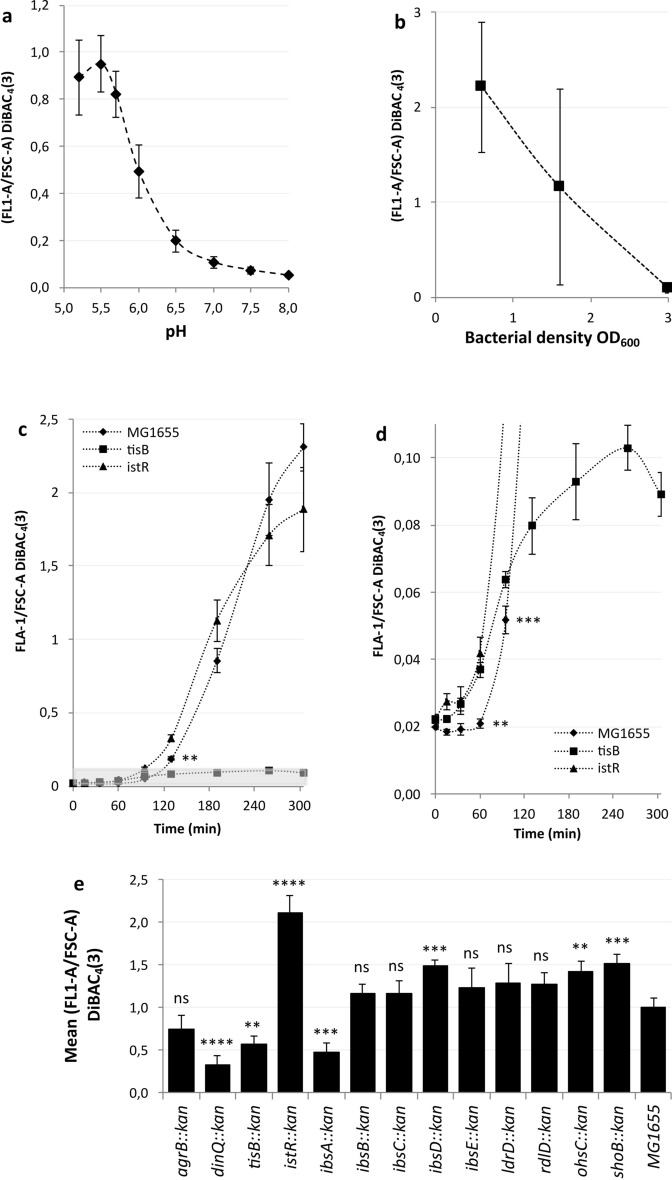
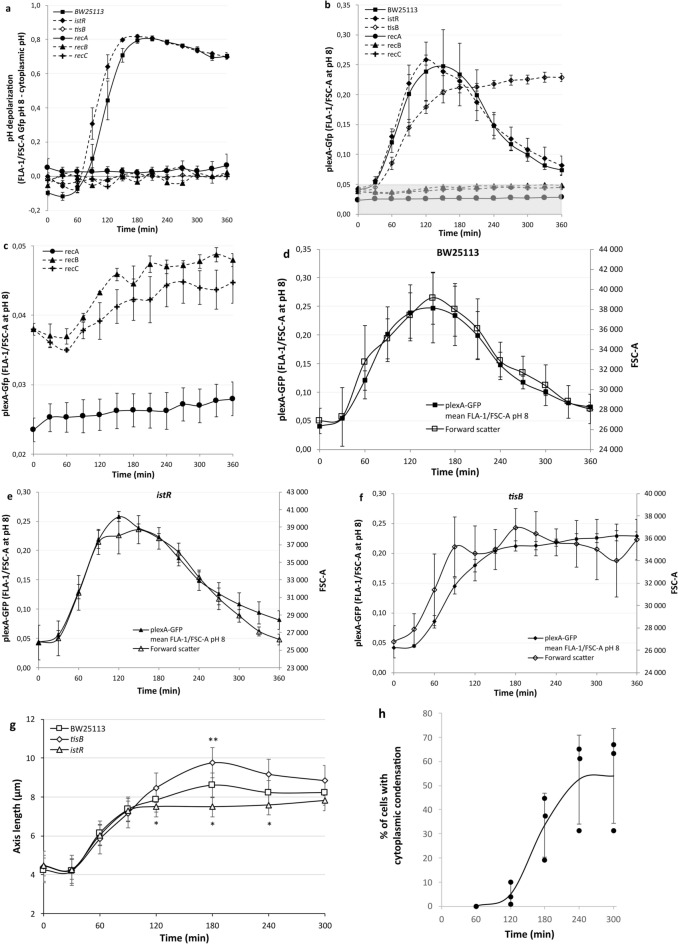
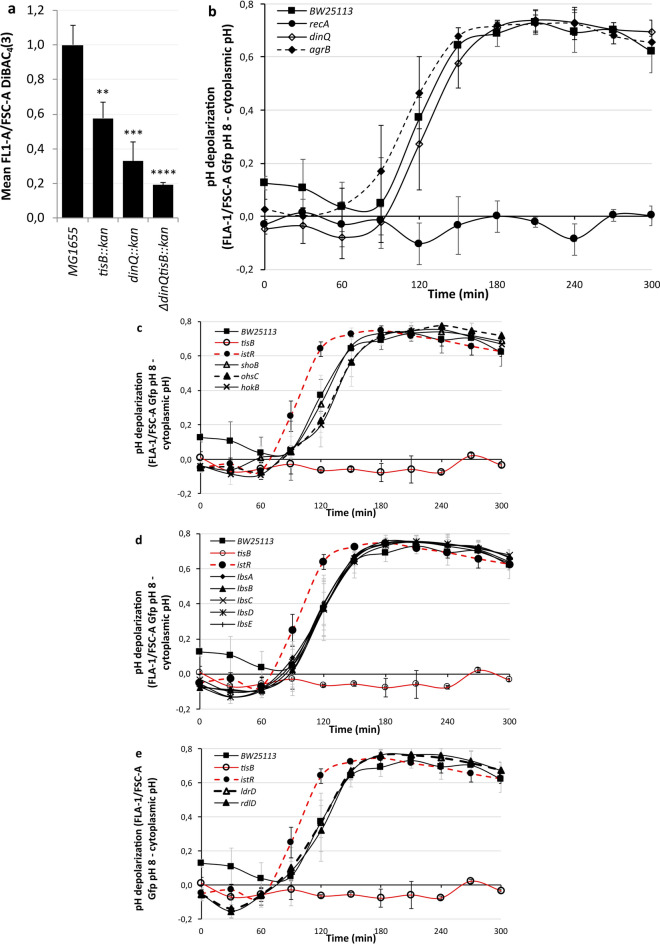
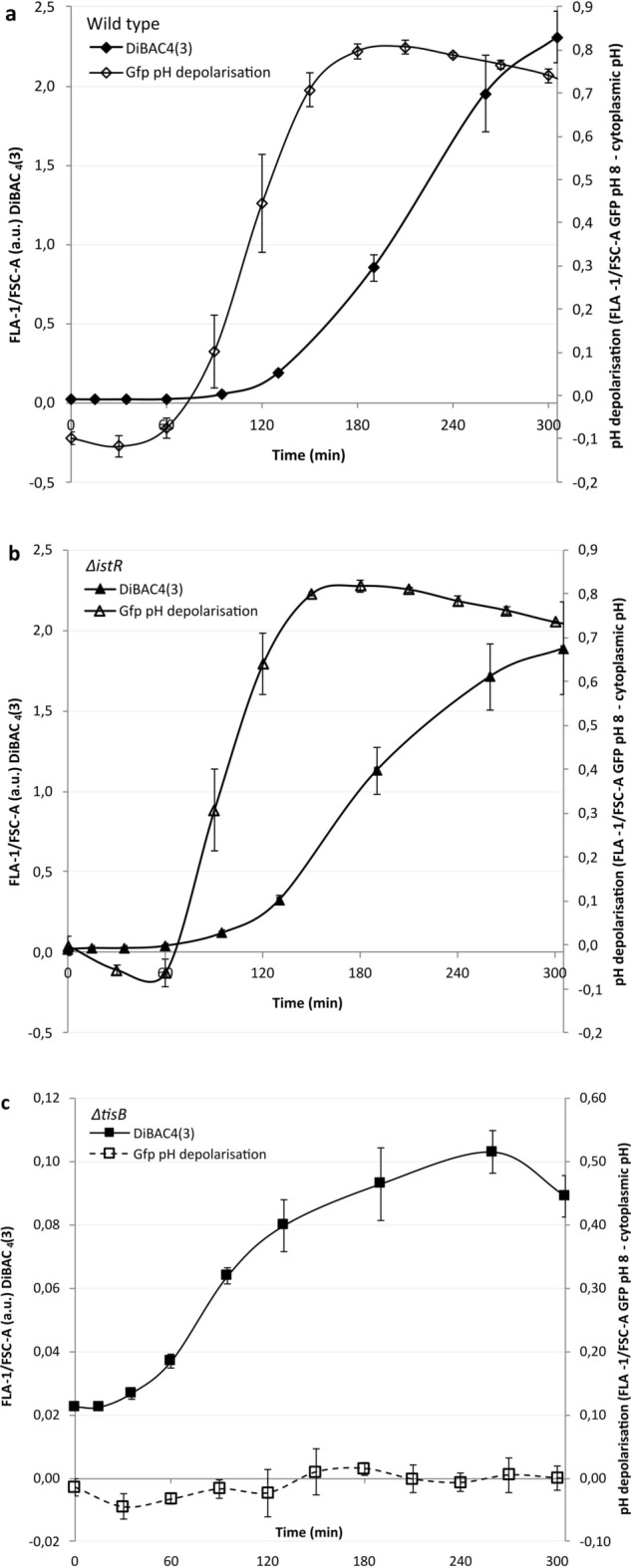
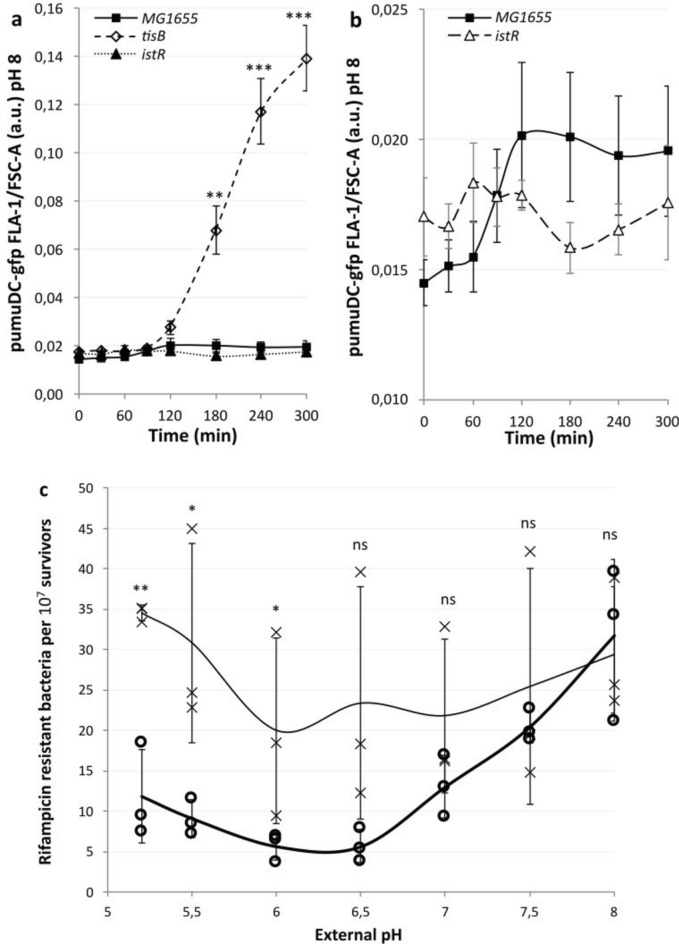
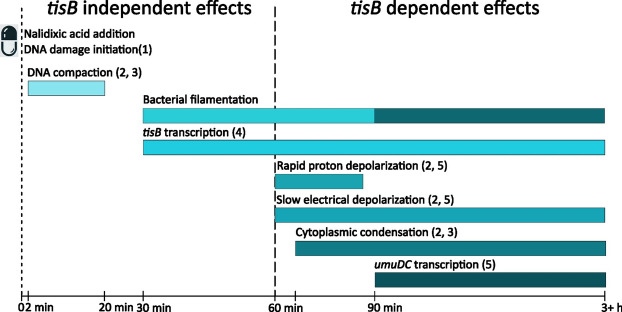
Bacterial toxin-antitoxin (TA) systems are genetic loci where the antitoxin gene product helps to control the expression or activity of the toxin gene product. Type I TA systems typically produce hydrophobic peptides that often localize to the inner membrane of bacteria. These amphipathic peptides can then potentially affect ion flows across the inner membrane. Here, we show that several type I toxins from Escherichia coli can affect depolarization, whereas tisB exclusively controls the depolarization of the proton gradient. tisB has been linked to persister cell formation following treatment with the antibiotic ciprofloxacin and tisB-istR has been implicated in the control of proton depolarization following treatment with ofloxacin. These results suggest that tisB could initiate the formation of persister cells by fully dissipating the proton gradient and that most of the electrical gradient greatly limiting ATP production following antibiotic-induced DNA damage.
Keywords: E. coli; Antibiotics; DNA damage; Membrane depolarization; TisB; Type I toxin antitoxin systems.
© 2025. The Author(s).
Conflict of interest statement
Delcaration. Competing interests: The authors declare that they have no competing interests.
Figures
References
-
- Kawano, M., Oshima, T., Kasai, H. & Mori, H. Molecular characterization of long direct repeat (LDR) sequences expressing a stable mRNA encoding for a 35-amino-acid cell-killing peptide and a cis-encoded small antisense RNA in escherichia coli. Mol. Microbiol.45, 333–349. 10.1046/j.1365-2958.2002.03042.x (2002). - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
