

A monomeric envelope glycoprotein cytoplasmic tail is sufficient for HIV-1 Gag lattice trapping and incorporation
- PMID: 40231821
- PMCID: PMC12090761
- DOI: 10.1128/jvi.02105-24
A monomeric envelope glycoprotein cytoplasmic tail is sufficient for HIV-1 Gag lattice trapping and incorporation
Abstract
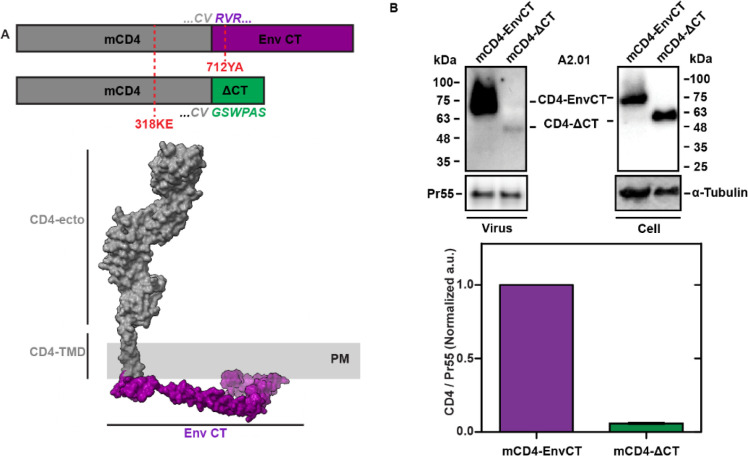
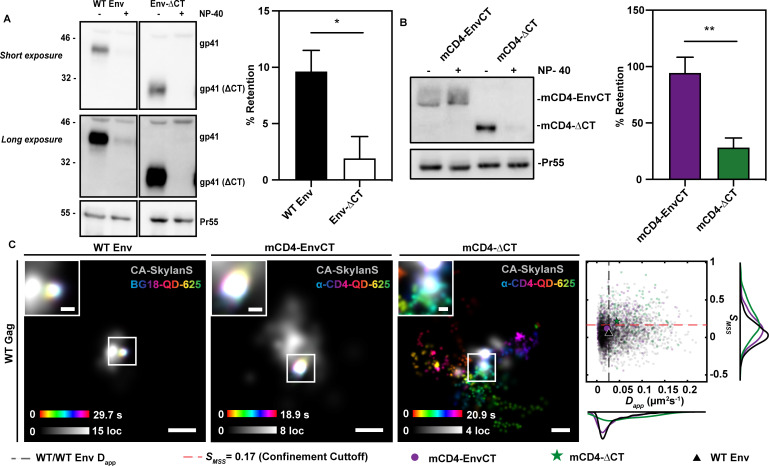
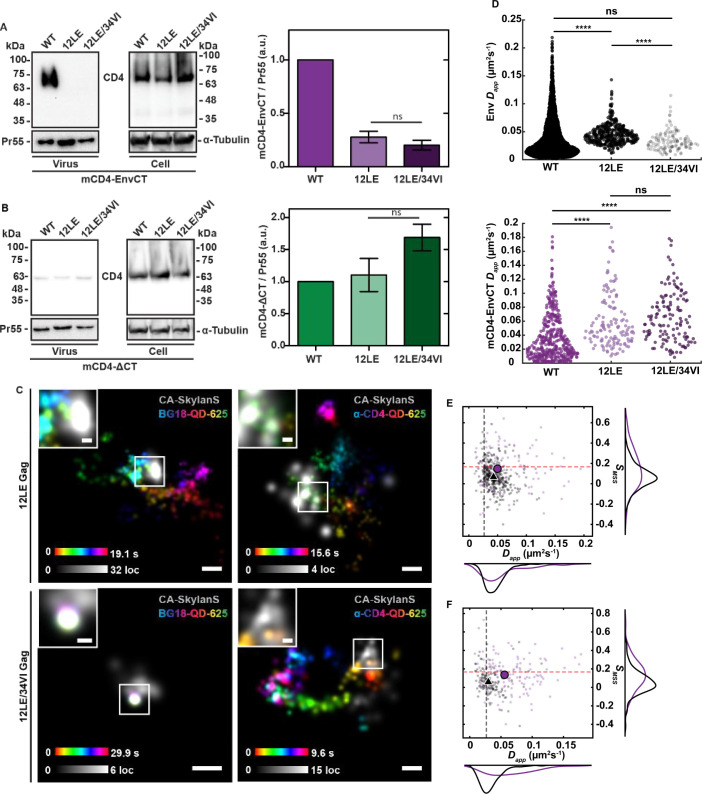
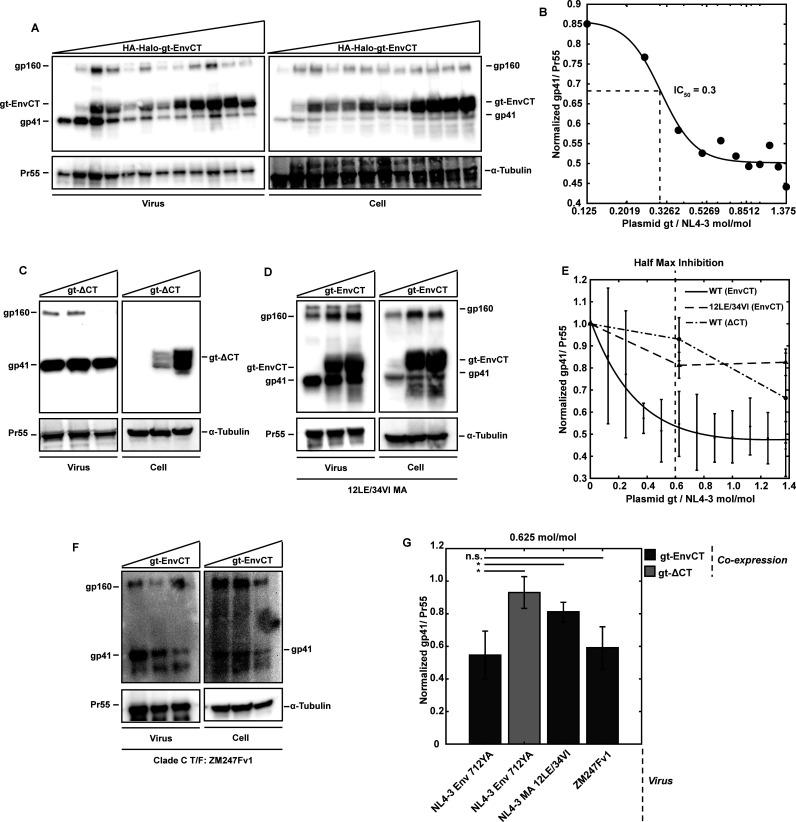
To become infectious, assembling enveloped viruses must acquire viral glycoproteins to mediate downstream infection events. Human immunodeficiency virus-1 (HIV-1) envelope glycoproteins (Env) are well characterized to function as trimers for membrane fusion and entry; however, we sought to understand whether the trimeric structure of Env is required for incorporation into virus particles. Using superresolution live-cell imaging and biochemical assays, we demonstrate that a monomeric receptor chimera containing the Env cytoplasmic tail (Env-CT), known to regulate Env incorporation, is sufficient for lattice trapping and incorporation into virus assembly sites. We also demonstrate that these Env-CT monomers can restrict the incorporation of native Env trimers, competing for an apparently limited number of interaction sites in each assembling particle. Furthermore, this monomeric construct can restrict the incorporation of Env glycoproteins from an evolutionarily distant HIV-1 primary isolate. Our findings support a model where a monomeric Env-CT mediates Env incorporation, with this mechanism of Env incorporation being conserved between distant clades of HIV-1.IMPORTANCETo combat the prevalence of HIV-1 and antiviral resistance, new classes of antivirals are needed. An attractive target for new classes includes virus assembly because released virus particles unable to obtain Env glycoproteins are non-infectious and unable to propagate HIV-1 infection. One requisite to the development of an antiviral targeting Gag-Env coalescence is the need to define the functional units constituting this molecular interface. Although Env functions as an obligatory trimer for virus entry, we demonstrate that a monomeric Env-CT is sufficient for Env incorporation into HIV-1 particles. Monomeric Env-CT displayed saturability in viral lattices and the ability to compete with native Env trimers for particle incorporation. These results suggest a less complex Env-CT structure mediates virus incorporation and that Env-CT mimetics could yield broad competitive activity against HIV-1 infection.
Keywords: Env; Gag; HIV-1; biochemistry; glycoprotein incorporation; single-particle tracking; virus assembly.
Conflict of interest statement
The authors declare no conflict of interest
Figures
Similar articles
-
HIV-1 Matrix Trimerization-Impaired Mutants Are Rescued by Matrix Substitutions That Enhance Envelope Glycoprotein Incorporation.J Virol. 2019 Dec 12;94(1):e01526-19. doi: 10.1128/JVI.01526-19. Print 2019 Dec 12. J Virol. 2019. PMID: 31619553 Free PMC article.
-
Single-molecule imaging of HIV-1 envelope glycoprotein dynamics and Gag lattice association exposes determinants responsible for virus incorporation.Proc Natl Acad Sci U S A. 2019 Dec 10;116(50):25269-25277. doi: 10.1073/pnas.1910008116. Epub 2019 Nov 22. Proc Natl Acad Sci U S A. 2019. PMID: 31757854 Free PMC article.
-
Trimer Enhancement Mutation Effects on HIV-1 Matrix Protein Binding Activities.J Virol. 2016 May 27;90(12):5657-5664. doi: 10.1128/JVI.00509-16. Print 2016 Jun 15. J Virol. 2016. PMID: 27030269 Free PMC article.
-
The Interplay between HIV-1 Gag Binding to the Plasma Membrane and Env Incorporation.Viruses. 2020 May 16;12(5):548. doi: 10.3390/v12050548. Viruses. 2020. PMID: 32429351 Free PMC article. Review.
-
The role of matrix in HIV-1 envelope glycoprotein incorporation.Trends Microbiol. 2014 Jul;22(7):372-8. doi: 10.1016/j.tim.2014.04.012. Epub 2014 Jun 2. Trends Microbiol. 2014. PMID: 24933691 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
