

HMGA1 deficiency: a pathogenic link between tau pathology and insulin resistance
- PMID: 40233659
- PMCID: PMC12019291
- DOI: 10.1016/j.ebiom.2025.105700
HMGA1 deficiency: a pathogenic link between tau pathology and insulin resistance
Abstract
Background: Growing evidence links tau-related neurodegeneration with insulin resistance and type 2 diabetes (T2D), though the underlying mechanisms remain unclear. Our previous research identified HMGA1 as crucial for insulin receptor (INSR) expression, with defects in the HMGA1 gene associated with insulin resistance and T2D. Here, we explore HMGA1 deficiency as a potential contributor to tauopathies, such as Alzheimer's disease (AD), and its connection to insulin resistance.
Methods: Immunoblot analyses, protein-DNA interaction studies, ChIP-qPCR, and reporter gene assays were conducted in human and mouse neuronal cell models. Tau immunohistochemistry, behavioural studies, and brain glucose metabolism were analysed in Hmga1-knockout mice. Additionally, a case-control study investigated the relationship between HMGA1 and tau pathology in patients with tauopathy, carrying or not the HMGA1 rs146052672 variant, known to reduce HMGA1 protein levels and increase the risk of insulin resistance and T2D.
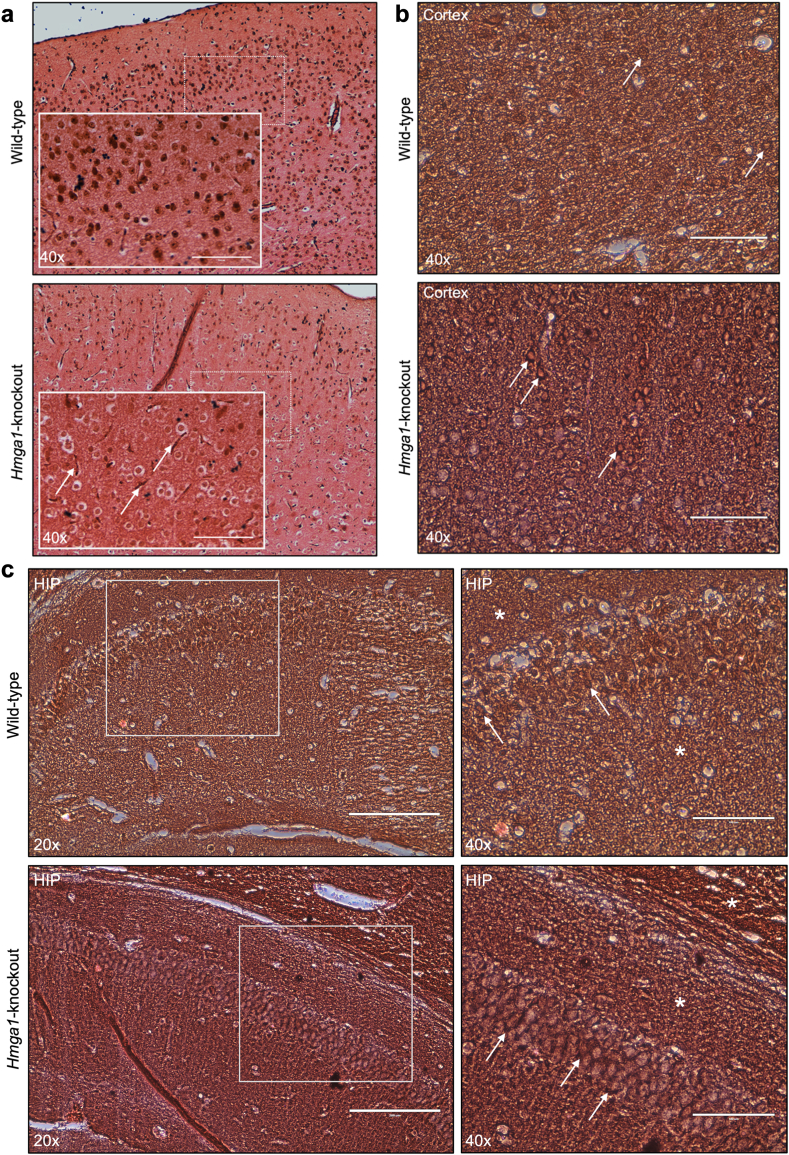
Findings: We show that HMGA1 regulates tau protein expression primarily through the specific repression of MAPT gene transcription. In both human neuronal cells and primary mouse neurons, tau mRNA and protein levels were inversely correlated with HMGA1 expression. This inverse relationship was further confirmed in the brain of Hmga1-knockout mice, where tau was overexpressed, INSR was downregulated, and brain glucose uptake was impaired. Additionally, the rs146052672 variant was more common in patients with tauopathy (12/69, 17.4%) than in controls (10/200, 5.0%) (p = 0.001), and carriers of this variant exhibited more severe disease progression and poorer therapeutic outcomes.
Interpretation: These findings suggest that HMGA1 deficiency may drive tau pathology, linking tauopathies to insulin resistance and providing new insights into the relationship between metabolic and neurodegenerative disorders. Furthermore, our observation that over 17% of individuals with tauopathy exhibit a deficit in HMGA1 protein production could have significant clinical implications, potentially guiding the development of therapeutic strategies targeting this specific defect.
Funding: See acknowledgements section.
Keywords: Gene transcription; Glucose metabolism; HMGA1; Insulin resistance; Neurodegeneration.
Copyright © 2025 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures









References
-
- Spillantini M.G., Schmidt M.L., Lee V.M., Trojanowski J.Q., Jakes R., Goedert M. Alpha-synuclein in Lewy bodies. Nature. 1997;388(6645):839–840. - PubMed
-
- Delacourte A. Tau pathology and neurodegeneration: an obvious but misunderstood link. J Alzheimers Dis. 2008;14(4):437–440. - PubMed
-
- Alonso A.D., Cohen L.S. Our tau tales from normal to pathological behavior. J Alzheimers Dis. 2018;64(s1):S507–S516. - PubMed
-
- Ballatore C., Lee V.M., Trojanowski J.Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 2007;8(9):663–672. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
