

METTL3 promotes an immunosuppressive microenvironment in bladder cancer via m6A-dependent CXCL5/CCL5 regulation
- PMID: 40234090
- PMCID: PMC12001370
- DOI: 10.1136/jitc-2024-011108
METTL3 promotes an immunosuppressive microenvironment in bladder cancer via m6A-dependent CXCL5/CCL5 regulation
Abstract
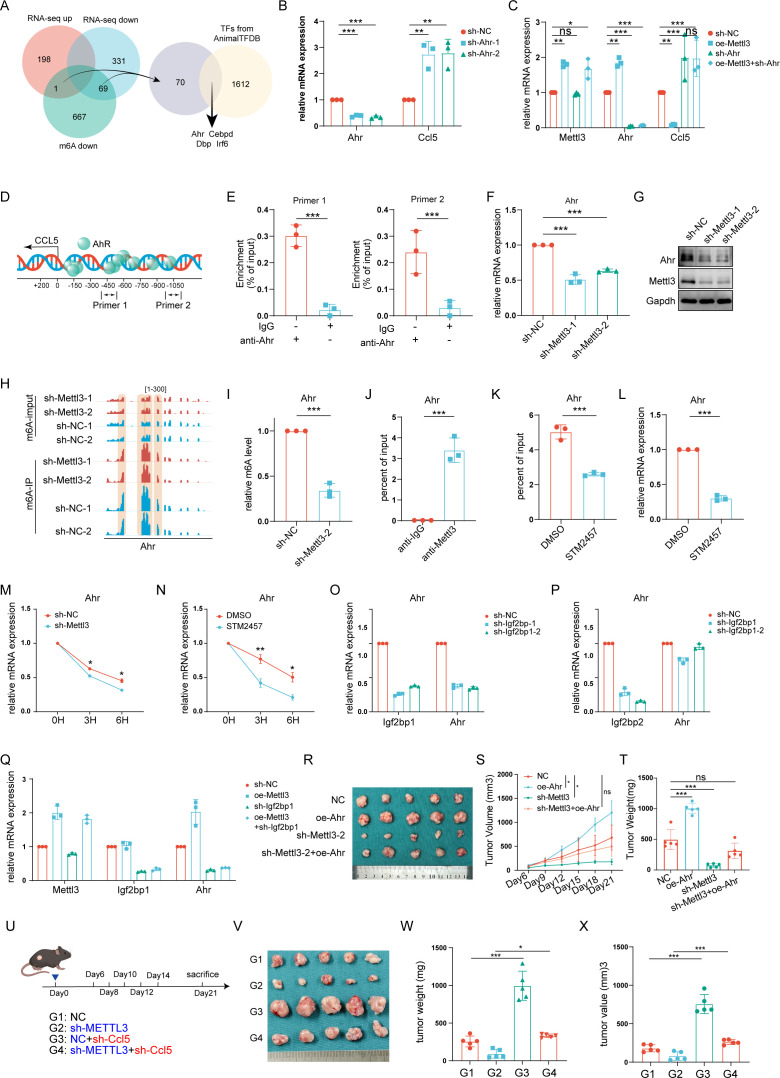
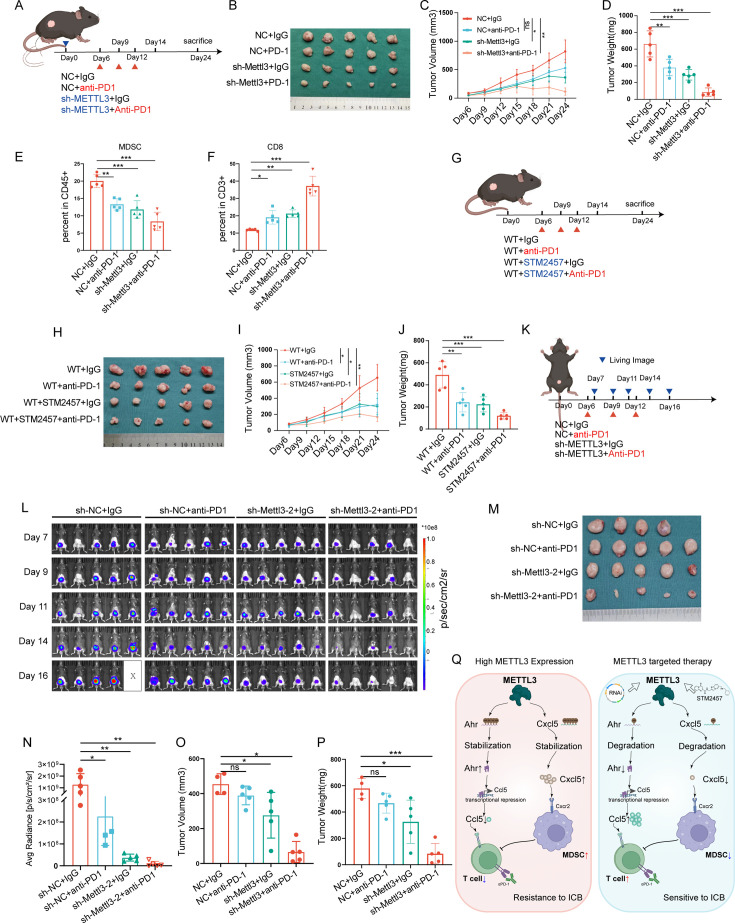
Background: Bladder cancer (BLCA) is a challenging malignancy with a poor prognosis, particularly in muscle-invasive cases. Despite recent advancements in immunotherapy, response rates remain suboptimal. This study investigates the role of METTL3, an m6A RNA methylation "writer," in regulating the immune microenvironment of BLCA.
Methods: Through bioinformatics analysis, we identified METTL3 as being associated with the formation of an immunosuppressive microenvironment in BLCA and poor response to immunotherapy. Subsequently, we silenced METTL3 expression in BLCA cells using short hairpin RNA (shRNA) or inhibited its function with STM2457. The effectiveness of these interventions in remodeling the BLCA tumor microenvironment (TME) was confirmed through animal experiments and flow cytometry. Mechanistically, RNA sequencing and methylated RNA immunoprecipitation (MeRIP) sequencing revealed the molecular pathways by which METTL3 regulates the TME. This was further validated using in vitro cell co-culture, immunoprecipitation, ELISA, and RNA degradation assays. The synergistic effect of METTL3 with anti-Programmed Cell Death Protein 1 (PD-1) treatment in BLCA was confirmed in both orthotopic and ectopic BLCA animal models.
Results: METTL3 was found to increase CXCL5 levels and suppress CCL5 expression in an m6A-dependent manner, leading to increased recruitment of myeloid-derived suppressor cells (MDSCs) and reduced infiltration of CD8+T cells. Silencing METTL3 or inhibiting its function restored immune cell balance and significantly enhanced the efficacy of anti-PD-1 therapy. Clinically, METTL3 overexpression correlated with poor complete response rate to immune checkpoint inhibitors (ICIs) therapy, associated with an immunosuppressive microenvironment characterized by elevated MDSC levels and reduced CD8+T cell infiltration.
Conclusions: These findings highlight METTL3 as a key regulator of the immune microenvironment in BLCA and a promising therapeutic target to improve immunotherapy outcomes. Targeting METTL3 could potentially enhance the efficacy of ICIs in patients with BLCA.
Keywords: Bladder Cancer; Cytokine; Immunotherapy; Myeloid-derived suppressor cell - MDSC; T cell.
© Author(s) (or their employer(s)) 2025. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ Group.
Conflict of interest statement
Competing interests: None declared.
Figures




Similar articles
-
TBX3 shapes an immunosuppressive microenvironment and induces immunotherapy resistance.Theranostics. 2025 Jan 6;15(5):1966-1986. doi: 10.7150/thno.103175. eCollection 2025. Theranostics. 2025. PMID: 39897553 Free PMC article.
-
METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner.Mol Cancer. 2019 Jun 22;18(1):110. doi: 10.1186/s12943-019-1036-9. Mol Cancer. 2019. PMID: 31228940 Free PMC article.
-
Targeting METTL3 as a checkpoint to enhance T cells for tumour immunotherapy.Clin Transl Med. 2024 Nov;14(11):e70089. doi: 10.1002/ctm2.70089. Clin Transl Med. 2024. PMID: 39568154 Free PMC article.
-
Myeloid-Derived Suppressor Cells in Bladder Cancer: An Emerging Target.Cells. 2024 Oct 27;13(21):1779. doi: 10.3390/cells13211779. Cells. 2024. PMID: 39513886 Free PMC article. Review.
-
The RNA m6A writer METTL3 in tumor microenvironment: emerging roles and therapeutic implications.Front Immunol. 2024 Jan 22;15:1335774. doi: 10.3389/fimmu.2024.1335774. eCollection 2024. Front Immunol. 2024. PMID: 38322265 Free PMC article. Review.
Cited by
-
Innovative Peptide Therapeutics in the Pipeline: Transforming Cancer Detection and Treatment.Int J Mol Sci. 2025 Jul 16;26(14):6815. doi: 10.3390/ijms26146815. Int J Mol Sci. 2025. PMID: 40725089 Free PMC article. Review.
-
Correspondence on "METTL3 promotes an immunosuppressive microenvironment in bladder cancer via m6A-dependent CXCL5/CCL5 regulation" by Tong et al.J Immunother Cancer. 2025 Jul 8;13(7):e012565. doi: 10.1136/jitc-2025-012565. J Immunother Cancer. 2025. PMID: 40633936 Free PMC article. No abstract available.
-
The impact of METTL3 on bladder cancer through m6A modification: a potential therapeutic target and prognostic biomarker.Front Oncol. 2025 Jul 3;15:1622117. doi: 10.3389/fonc.2025.1622117. eCollection 2025. Front Oncol. 2025. PMID: 40678060 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials