

Functionally distinct GABAergic amacrine cell types regulate spatiotemporal encoding in the mouse retina
- PMID: 40234708
- PMCID: PMC12148929
- DOI: 10.1038/s41593-025-01935-0
Functionally distinct GABAergic amacrine cell types regulate spatiotemporal encoding in the mouse retina
Abstract
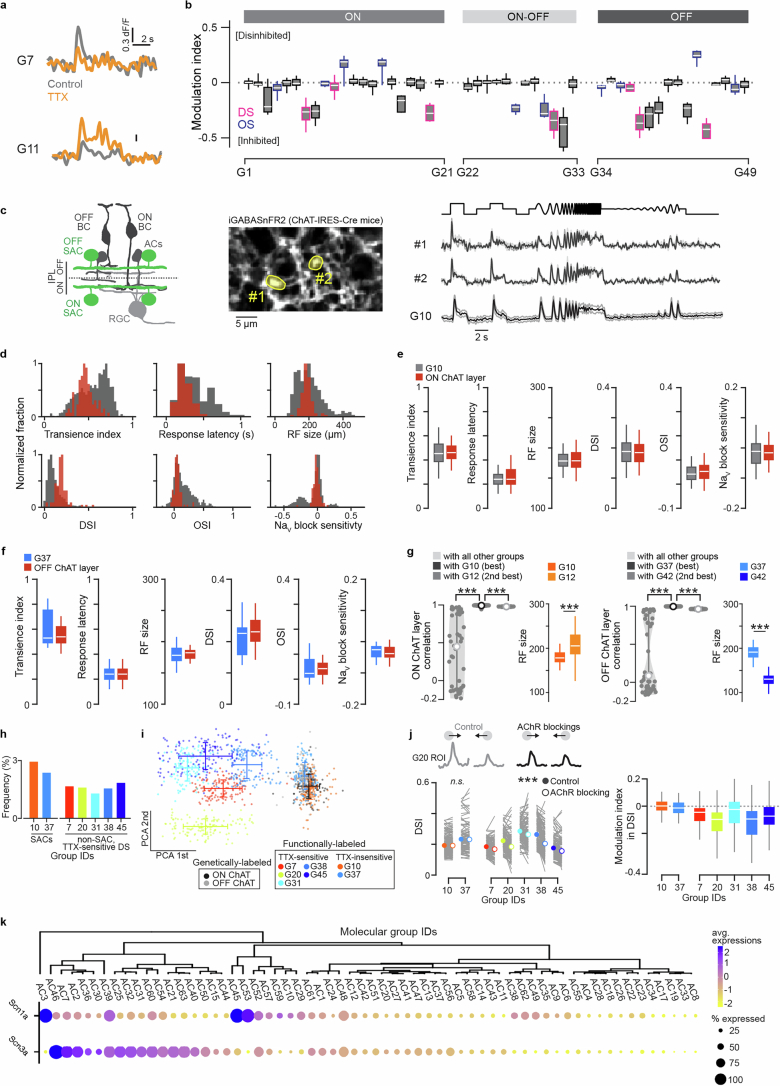
GABA (γ-aminobutyric acid) is the primary inhibitory neurotransmitter in the mammalian central nervous system. GABAergic neuronal types play important roles in neural processing and the etiology of neurological disorders; however, there is no comprehensive understanding of their functional diversity. Here we perform two-photon imaging of GABA release in the inner plexiform layer of male and female mice retinae (8-16 weeks old) using the GABA sensor iGABASnFR2. By applying varied light stimuli to isolated retinae, we reveal over 40 different GABA-releasing neuron types. Individual types show layer-specific visual encoding within inner plexiform layer sublayers. Synaptic input and output sites are aligned along specific retinal orientations. The combination of cell type-specific spatial structure and unique release kinetics enables inhibitory neurons to sculpt excitatory signals in response to a wide range of behaviorally relevant motion structures. Our findings emphasize the importance of functional diversity and intricate specialization of GABAergic neurons in the central nervous system.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures














References
MeSH terms
Substances
Grants and funding
- R344-2020-300/Lundbeckfonden (Lundbeck Foundation)
- R248-2016-2518/Lundbeckfonden (Lundbeck Foundation)
- NNF15OC0017252/Novo Nordisk Fonden (Novo Nordisk Foundation)
- NNF20OC0064395/Novo Nordisk Fonden (Novo Nordisk Foundation)
- 638730/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 20K23377/MEXT | Japan Society for the Promotion of Science (JSPS)
- 22K21353/MEXT | Japan Society for the Promotion of Science (JSPS)
- 23H04241/MEXT | Japan Society for the Promotion of Science (JSPS)
- 24H02311/MEXT | Japan Society for the Promotion of Science (JSPS)
- 23K19412/MEXT | Japan Society for the Promotion of Science (JSPS)
- JPMJPR2489/MEXT | Japan Society for the Promotion of Science (JSPS)
- 27786/Velux Fonden (Velux Foundation)
LinkOut - more resources
Full Text Sources
