

Transcriptional Intermediary Factor 1γ-Induced Irisin in Skeletal Muscle Attenuates Renal Fibrosis in Diabetic Nephropathy
- PMID: 40235186
- PMCID: PMC12000539
- DOI: 10.1002/jcsm.13810
Transcriptional Intermediary Factor 1γ-Induced Irisin in Skeletal Muscle Attenuates Renal Fibrosis in Diabetic Nephropathy
Abstract
Background: Transcriptional intermediary factor 1γ (TIF1γ) is a negative regulator of TGF-β1 signalling and has been associated with patient survival in renal cell carcinoma. However, its role in diabetes mellitus (DM), particularly in diabetic nephropathy (DN), remains unclear. DN is the leading cause of chronic kidney disease (CKD). We investigated the potential role of TIF1γ in mitigating multiple DM-related complications.
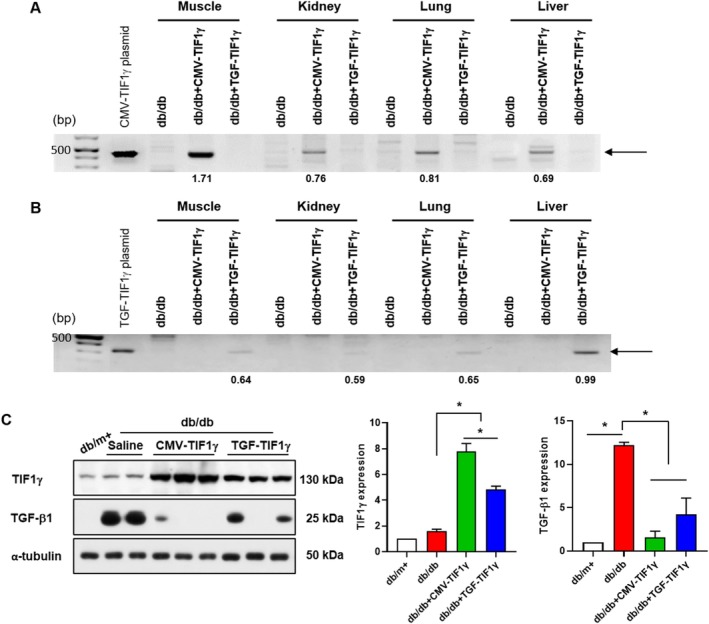
Methods: Mice were divided into four groups: db/m+, db/db and db/db mice treated with cytomegalovirus- or TGF-TIF1γ plasmids (40 μg/mouse; intraperitoneally weekly for 16 weeks). Renal injury, fibrosis, function and gene expression related to fibrosis and epithelial-mesenchymal transition (EMT) in the kidneys were assessed. Muscle atrophy, regeneration markers, myokine levels and exercise capacity were evaluated. C2C12 cells were exposed to palmitate with or without TIF1γ transfection, and irisin expression and secretion were measured. Muscle-kidney crosstalk was analysed using conditioned media (CM) from TIF1γ-transfected C2C12 cells in palmitate-treated human kidney (HK)-2 cells. Additionally, HK-2 cells were incubated in CM from fibronectin type III domain-containing protein (FNDC)5-knockdown C2C12 cells to confirm irisin-mediated kidney crosstalk by TIF1γ.
Results: TIF1γ treatment in db/db mice resulted in a significant attenuation of renal tubulointerstitial fibrosis (1.5-fold decrease), glomerular injury (1.8-fold improvement), tubular injury (1.6-fold improvement), renal dysfunction (1.7-fold improvement) and a reduction in EMT-related factors (1.8-fold decrease) (p < 0.05). The levels of administered TIF1γ plasmids were higher in skeletal muscle than in renal tissues. TIF1γ expression was significantly elevated in the skeletal muscle of db/db mice treated with TIF1γ plasmids (6.5-fold) (p < 0.05). Mice receiving both plasmids exhibited a 1.8-fold reduction in pathological muscle morphology and atrophy-related gene expression, a 3.0-fold increase in regeneration-related gene expression and a 1.6-fold improvement in muscle function (p < 0.05). Irisin expression increased by 2.1-fold in skeletal muscle and serum (p < 0.05). In TIF1γ-transfected C2C12 cells, irisin secretion was elevated by 1.5-fold (p < 0.05). CM from TIF1γ-transfected C2C12 cells attenuated EMT in palmitate-treated HK-2 cells, compared with medium from nontransfected C2C12 cells (1.9-fold improvement [p < 0.05]). Conversely, FNDC5 knockdown in C2C12 cells accelerated EMT in palmitate-treated HK-2 cells, as evidenced by decreased bone morphogenetic protein-7 (1.6-fold) and increased EMT-related factors (2.1-fold) (p < 0.05), compared with palmitate alone and small interfering RNA control.
Conclusions: Our findings emphasize the potential of TIF1γ as a multitargeted therapeutic agent for DN, mitigating both renal and muscular complications through direct fibrosis inhibition and indirect myokine-mediated inter-organ crosstalk.
Keywords: chronic kidney disease; diabetic nephropathy; fibrosis; irisin; muscle‐kidney crosstalk; transcriptional intermediary factor 1γ.
© 2025 The Author(s). Journal of Cachexia, Sarcopenia and Muscle published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- McClellan W. M. and Flanders W. D., “Risk Factors for Progressive Chronic Kidney Disease,” Journal of the American Society of Nephrology 14 (2003): S65–S70. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
