

Chloride intracellular channel (CLIC) protein function in S1P-induced Rac1 activation requires membrane localization of the C-terminus, but not thiol-transferase nor ion channel activities
- PMID: 40235733
- PMCID: PMC11996907
- DOI: 10.3389/fcell.2025.1565262
Chloride intracellular channel (CLIC) protein function in S1P-induced Rac1 activation requires membrane localization of the C-terminus, but not thiol-transferase nor ion channel activities
Abstract
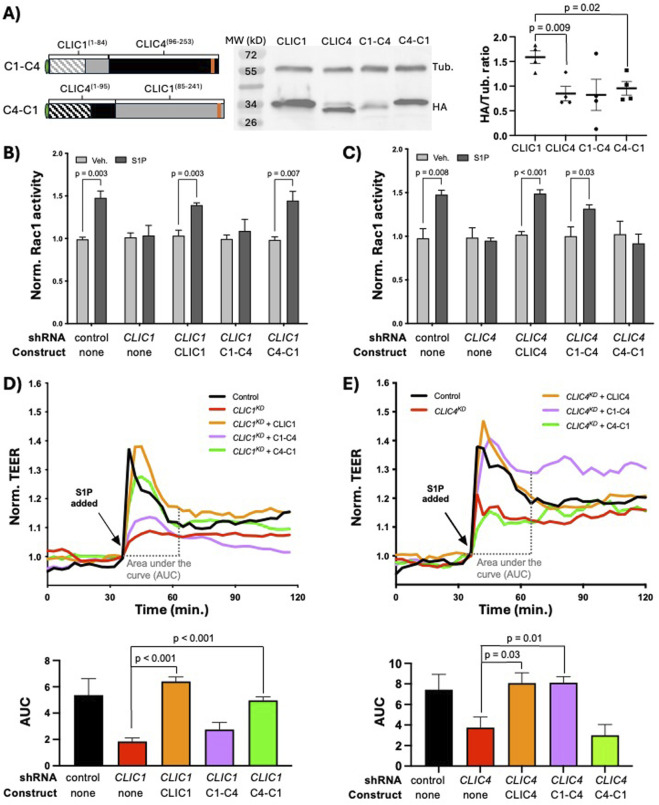
We have established a novel and evolutionarily-conserved function for chloride intracellular channel proteins (CLICs) in regulating Rho/Rac GTPases downstream of G protein-coupled receptors (GPCRs). Endothelial CLIC1 and CLIC4 are rapidly and transiently re-localized from the cytoplasm to the plasma membrane in response to the GPCR ligand sphingosine-1-phosphate (S1P), and both CLICs are required to activate Rac1 in response to S1P, but how they perform this function remains unknown. Biochemical studies suggest that CLICs act as non-specific ion channels and/or as glutathione-S-transferases, dependent on N-terminal features, in vitro. Here we investigate CLIC functional domains and membrane localization requirements for their function in S1P-mediated Rac1 signaling. Structure-function analyses of CLIC function in endothelial cells demonstrate that CLIC1 and CLIC4-specific functions reside at their C-termini, and that the CLIC4 N-terminus encodes determinants required for S1P-induced re-localization to the plasma membrane but is dispensable for S1P-induced Rac1 activation when the C-terminus is localized to the plasma membrane via a heterologous signal. Our results demonstrate that the postulated ion channel and thiol-transferase activities of CLICs are not required for Rac1 activation and suggests that sequences in the CLIC C-termini are critical for this function. Given the importance of S1P signaling in vascular biology and disease, our work establishes a platform to further our understanding of the membrane-localized proteins required to link GPCR activity to Rho/Rac regulation.
Keywords: CLIC1; CLIC4; Rac1; RhoA; S1P receptor; endothelium; sphingosine-1-phosphate.
Copyright © 2025 Mao, Jesse, Shaye and Kitajewski.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Update of
-
Chloride intracellular channel (CLIC) protein function in S1P-induced Rac1 activation requires membrane localization of the C-terminus, but not thiol-transferase nor ion channel activities.bioRxiv [Preprint]. 2025 Jan 24:2025.01.22.634370. doi: 10.1101/2025.01.22.634370. bioRxiv. 2025. Update in: Front Cell Dev Biol. 2025 Apr 01;13:1565262. doi: 10.3389/fcell.2025.1565262. PMID: 39896666 Free PMC article. Updated. Preprint.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous