

Molecular structure and enzymatic mechanism of the human collagen hydroxylysine galactosyltransferase GLT25D1/COLGALT1
- PMID: 40240392
- PMCID: PMC12003778
- DOI: 10.1038/s41467-025-59017-5
Molecular structure and enzymatic mechanism of the human collagen hydroxylysine galactosyltransferase GLT25D1/COLGALT1
Abstract
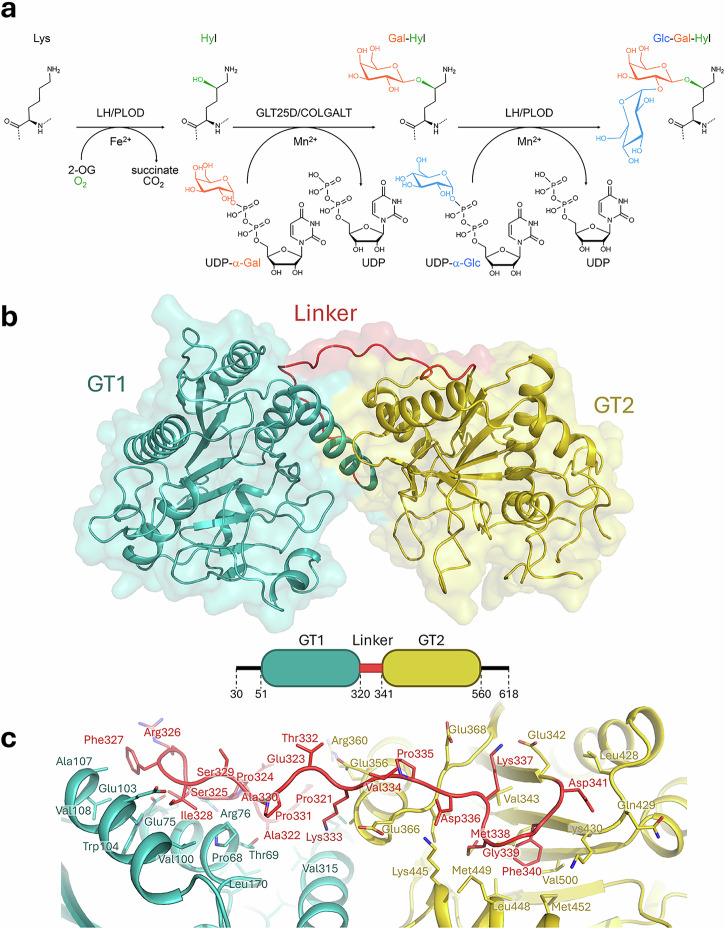
During collagen biosynthesis, lysine residues undergo extensive post-translational modifications through the alternate action of two distinct metal ion-dependent enzyme families (i.e., LH/PLODs and GLT25D/COLGALT), ultimately producing the highly conserved α-(1,2)-glucosyl-β-(1,O)-galactosyl-5-hydroxylysine pattern. Malfunctions in these enzymes are linked to developmental pathologies and extracellular matrix alterations associated to enhanced aggressiveness of solid tumors. Here, we characterized human GLT25D1/COLGALT1, revealing an elongated head-to-head homodimeric assembly. Each monomer encompasses two domains (named GT1 and GT2), both unexpectedly capable of binding metal ion cofactors and UDP-α-galactose donor substrates, resulting in four candidate catalytic sites per dimer. We identify the catalytic site in GT2, featuring an unusual Glu-Asp-Asp motif critical for Mn2+ binding, ruling out direct catalytic roles for the GT1 domain, but showing that in this domain the unexpectedly bound Ca2+ and UDP-α-galactose cofactors are critical for folding stability. Dimerization, albeit not essential for GLT25D1/COLGALT1 activity, provides a critical molecular contact site for multi-enzyme assembly interactions with partner multifunctional LH/PLOD lysyl hydroxylase-glycosyltransferase enzymes.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures





References
-
- Myllyharju, J. & Kivirikko, K. I. Collagens and collagen-related diseases. Ann. Med.33, 7–21 (2001). - PubMed
-
- Ishikawa, Y. & Bachinger, H. P. A molecular ensemble in the rER for procollagen maturation. Biochim. Biophys. Acta1833, 2479–2491 (2013). - PubMed
-
- Gelse, K., Poschl, E. & Aigner, T. Collagens-structure, function, and biosynthesis. Adv. Drug Deliv. Rev.55, 1531–1546 (2003). - PubMed
MeSH terms
Substances
Grants and funding
- MFAG 20075, BRIDGE 27004/Associazione Italiana per la Ricerca sul Cancro (Italian Association for Cancer Research)
- Rarer Types EDS Grant 2022/Ehlers-Danlos Society (EDS)
- CDA 2013/Giovanni Armenise-Harvard Foundation
- NextGeneration-EU PNRR MUR M4C2 PE00000007 INF-ACT/Ministero dell'Istruzione, dell'Università e della Ricerca (Ministry of Education, University and Research)
- PRIN PNRR 2022 P20224WAME/Ministero dell'Istruzione, dell'Università e della Ricerca (Ministry of Education, University and Research)
- PRIN PNRR 2022 P20224WAME/Ministero dell'Istruzione, dell'Università e della Ricerca (Ministry of Education, University and Research)
- Piano Operativo Salute, IMMUNO-HUB/Ministero della Salute (Ministry of Health, Italy)
- regional law n° 9/2020, resolution n° 3776/2020/Regione Lombardia (Region of Lombardy)
LinkOut - more resources
Full Text Sources
Miscellaneous
