

Functional Analysis of Mannosyltransferase-Related Genes UvALGs in Ustilaginoidea virens
- PMID: 40243604
- PMCID: PMC11988645
- DOI: 10.3390/ijms26072979
Functional Analysis of Mannosyltransferase-Related Genes UvALGs in Ustilaginoidea virens
Abstract
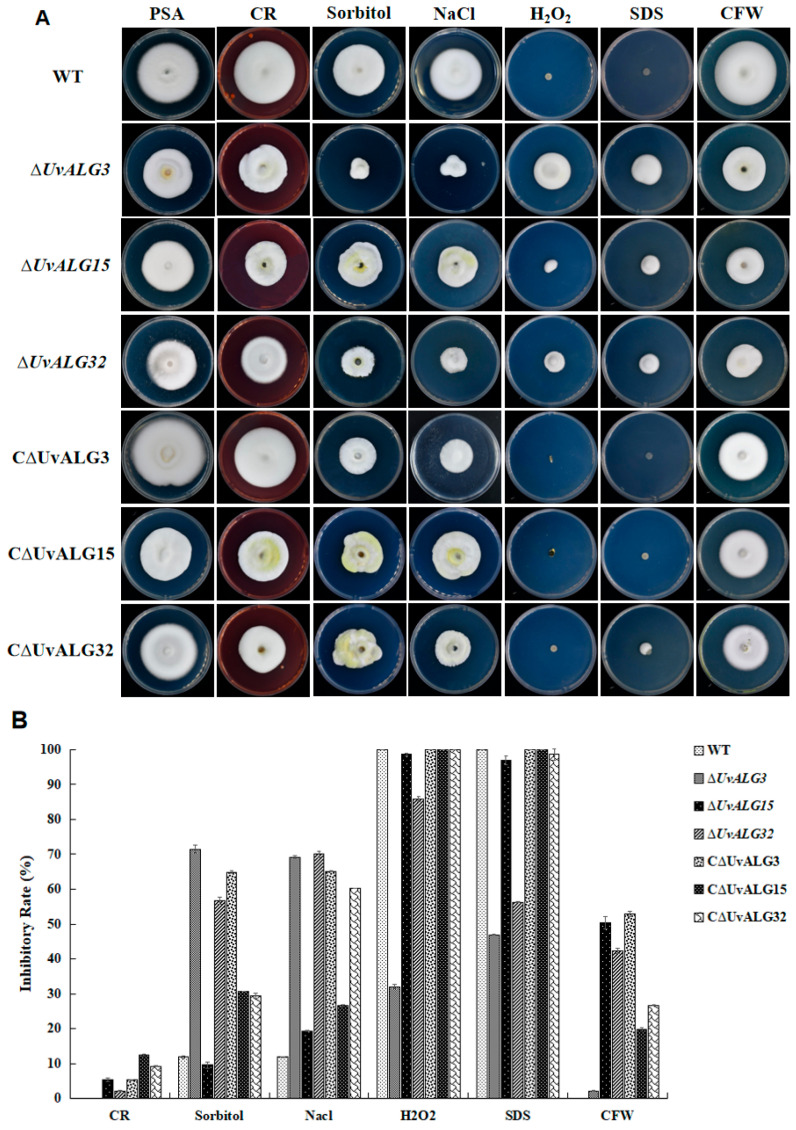
Rice false smut, caused by Ustilaginoidea virens, is one of the three major rice diseases in China. It not only seriously affects the rice yield and quality but also endangers human and animal health. Studying the pathogenic mechanism of U. virens has important theoretical significance and application value for clarifying the infection characteristics of the pathogen and cultivating disease-resistant varieties. Plant pathogenic fungi utilize secreted effectors to suppress plant immune responses, which can function in the apoplast or within host cells and are likely glycosylated. However, the posttranslational regulation of these effectors remains unexplored. Deletion of ΔUvALG led to the cessation of secondary infection hyphae growth and a notable decrease in virulence. We observed that ΔUvALG mutants triggered a significant increase in reactive species production within host cells, akin to ALG mutants, which plays a crucial role in halting the growth of infection hyphae in the mutants. ALG functions by sequestering chitin oligosaccharides to prevent their recognition by the rice chitin elicitor, thereby inhibiting the activation of innate immune responses, including reactive species production. Our findings reveal that ALG3 possesses three N-glycosylation sites, and the simultaneous Alg-mediated N-glycosylation of each site is essential for maintaining protein stability and chitin-binding activity, both of which are critical for its effector function. These outcomes underscore the necessity of the Alg-mediated N-glycosylation of ALG to evade host innate immunity.
Keywords: Agrobacterium tumefaciens-mediated transformation; Ustilaginoidea virens; pathogenicity; target gene knockout.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







Similar articles
-
Rice false smut virulence protein subverts host chitin perception and signaling at lemma and palea for floral infection.Plant Cell. 2024 May 1;36(5):2000-2020. doi: 10.1093/plcell/koae027. Plant Cell. 2024. PMID: 38299379 Free PMC article.
-
N-glycosylation of effector proteins by an α-1,3-mannosyltransferase is required for the rice blast fungus to evade host innate immunity.Plant Cell. 2014 Mar;26(3):1360-76. doi: 10.1105/tpc.114.123588. Epub 2014 Mar 18. Plant Cell. 2014. PMID: 24642938 Free PMC article.
-
The Conserved Effector UvHrip1 interacts with OsHGW, and Infection of Ustilaginoidea virens Regulates Defense- and Heading Date-Related Signaling Pathway.Int J Mol Sci. 2020 May 10;21(9):3376. doi: 10.3390/ijms21093376. Int J Mol Sci. 2020. PMID: 32397668 Free PMC article.
-
Ustilaginoidea virens, an emerging pathogen of rice: the dynamic interplay between the pathogen virulence strategies and host defense.Planta. 2024 Sep 11;260(4):92. doi: 10.1007/s00425-024-04523-x. Planta. 2024. PMID: 39261328 Review.
-
Current understanding on Villosiclava virens, a unique flower-infecting fungus causing rice false smut disease.Mol Plant Pathol. 2016 Dec;17(9):1321-1330. doi: 10.1111/mpp.12362. Epub 2016 Apr 13. Mol Plant Pathol. 2016. PMID: 26720072 Free PMC article. Review.
References
-
- Andargie M., Li J. Expression of the Arabidopsis SWEET genes during rice false smut infection in the transgenic Arabidopsis thaliana containing increased levels of ATP and sucrose. Curr. Plant Biol. 2019;28:509–520. doi: 10.1007/s13562-019-00512-4. - DOI
-
- Andargie M., Li L.Y., Feng A.Q., Zhu X.Y., Li J.X. Development of a GFP-expressing Ustilaginoidea virens strain to study fungal invasion and colonization in rice spikelets. S. Afr. J. Bot. 2015;97:16–24. doi: 10.1016/j.sajb.2014.11.013. - DOI
-
- Ladhalakshmi D., Srinivas Prasad M., Prakasam V., Krishnaveni D., Sailaja B., Ram S., Vindeswari P., Jagjeet Singh L., Jyoti J., Surendran M., et al. Geographic distribution of false smut disease of rice in India and efficacy of selected fungicides for its management. Int. J. Pest Manag. 2019;65:177–185. doi: 10.1080/09670874.2018.1494865. - DOI
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
