

The Proline Dehydrogenase Gene CsProDH1 Regulates Homeostasis of the Pro-P5C Cycle Under Drought Stress in Tea Plants
- PMID: 40243904
- PMCID: PMC11988676
- DOI: 10.3390/ijms26073121
The Proline Dehydrogenase Gene CsProDH1 Regulates Homeostasis of the Pro-P5C Cycle Under Drought Stress in Tea Plants
Abstract
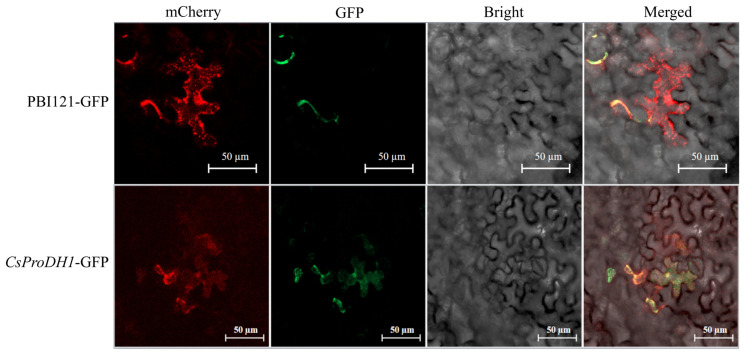
The homeostasis of the proline-Δ1-pyrroline-5-carboxylate (Pro-P5C) cycle, mediated by proline dehydrogenase (ProDH), plays a critical role in plants in response to abiotic stresses. The biological function of gene CsProDH1 under drought stress and its effects on amino acid metabolism and photosynthesis through proline metabolism were investigated. Enzymatic characterization of the CsProDH1 protein was conducted in vitro. Overexpression of CsProDH1 aggravated plant stress, as evident by reduced photosynthetic efficiency and increased reactive oxygen species, which activated the Pro-P5C cycle. In contrast, silencing CsProDH1 enhanced plant drought resistance, increased proline accumulation, and protected photosynthesis. Studies indicate that exogenous amino acid application mitigates drought-induced physiological impairments in plants by maintaining cellular homeostasis, with particular efficacy observed in enhancing tea plant drought resilience through improved osmotic adjustment and antioxidant capacity. This study uncovers the significant role of CsProDH1 in plant drought resistance and its regulatory mechanism, offering potential gene targets and application strategies for enhancing crop drought resistance.
Keywords: Pro-P5C cycle; drought stress; proline dehydrogenase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Unraveling delta1-pyrroline-5-carboxylate-proline cycle in plants by uncoupled expression of proline oxidation enzymes.J Biol Chem. 2009 Sep 25;284(39):26482-92. doi: 10.1074/jbc.M109.009340. Epub 2009 Jul 27. J Biol Chem. 2009. PMID: 19635803 Free PMC article.
-
Proline dehydrogenase contributes to pathogen defense in Arabidopsis.Plant Physiol. 2011 Apr;155(4):1947-59. doi: 10.1104/pp.110.167163. Epub 2011 Feb 10. Plant Physiol. 2011. PMID: 21311034 Free PMC article.
-
Context of action of proline dehydrogenase (ProDH) in the Hypersensitive Response of Arabidopsis.BMC Plant Biol. 2014 Jan 13;14:21. doi: 10.1186/1471-2229-14-21. BMC Plant Biol. 2014. PMID: 24410747 Free PMC article.
-
Regulation of levels of proline as an osmolyte in plants under water stress.Plant Cell Physiol. 1997 Oct;38(10):1095-102. doi: 10.1093/oxfordjournals.pcp.a029093. Plant Cell Physiol. 1997. PMID: 9399433 Review.
-
Proline dehydrogenase (oxidase) in cancer.Biofactors. 2012 Nov-Dec;38(6):398-406. doi: 10.1002/biof.1036. Epub 2012 Aug 8. Biofactors. 2012. PMID: 22886911 Free PMC article. Review.
References
-
- Yang X., Lu M., Wang Y., Wang Y., Liu Z., Chen S. Response Mechanism of Plants to Drought Stress. Horticulturae. 2021;7:50. doi: 10.3390/horticulturae7030050. - DOI
-
- Wang Z., Yang Y., Yadav V., Zhao W., He Y., Zhang X., Wei C. Drought-Induced Proline Is Mainly Synthesized in Leaves and Transported to Roots in Watermelon under Water Deficit. Hortic. Plant J. 2022;8:615–626. doi: 10.1016/j.hpj.2022.06.009. - DOI
-
- Machado J., Fernandes A.P.G., Bokor B., Vaculík M., Kostoláni D., Kokavcová A., Heuvelink E., Vasconcelos M.W., Carvalho S.M.P. Tomato Responses to Nitrogen, Drought and Combined Stresses: Shared and Specific Effects on Vascular Plant Anatomy, Nutrient Partitioning and Amino Acids Profile. Plant Physiol. Biochem. 2025;221:109649. doi: 10.1016/j.plaphy.2025.109649. - DOI - PubMed
-
- Ashraf M., Foolad M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007;59:206–216. doi: 10.1016/j.envexpbot.2005.12.006. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
