

Combined metabolic and enzymatic engineering for de novo biosynthesis of δ-tocotrienol in Yarrowia lipolytica
- PMID: 40248488
- PMCID: PMC12002712
- DOI: 10.1016/j.synbio.2025.02.011
Combined metabolic and enzymatic engineering for de novo biosynthesis of δ-tocotrienol in Yarrowia lipolytica
Abstract
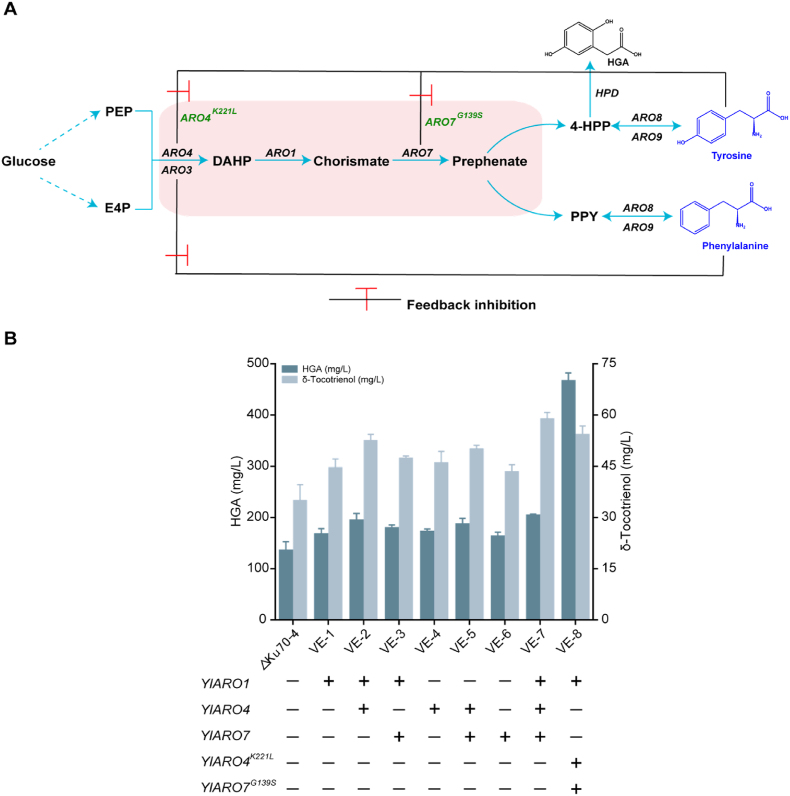
δ-Tocotrienol, an isomer of vitamin E with anti-inflammatory, neuroprotective and anti-coronary arteriosclerosis properties, is widely used in health care, medicine and other fields. Microbial synthesis of δ-tocotrienol offers significant advantages over plant extraction and chemical synthesis methods, including increased efficiency, cost-effectiveness and environmental sustainability. However, limited precursor availability and low catalytic efficiency of key enzymes remain major bottlenecks in the biosynthesis of δ-tocotrienol. In this study, we assembled the complete δ-tocotrienol biosynthetic pathway in Yarrowia lipolytica and enhanced the precursor supply, resulting in a titre of 102.8 mg/L. The catalytic efficiency of the rate-limiting steps in the pathway was then enhanced through various strategies, including fusion expression of key enzymes homogentisate phytyltransferaseand and tocopherol cyclase, semi-rational design of SyHPT and multi-copy integration of pathway genes. The final a δ-tocotrienol titre in a 5 L bioreactor was 466.8 mg/L following fed-batchfermentation. This study represents the first successful de novo biosynthesis of δ-tocotrienol in Y. lipolytica, providing valuable insights into the synthesis of vitamin E-related compounds.
Keywords: Fed-batch fermentation; Metabolic engineering; Yarrowia lipolytica; enzyme engineering; δ-tocotrienol.
© 2025 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures







Similar articles
-
De Novo High-Titer Production of Delta-Tocotrienol in Recombinant Saccharomyces cerevisiae.J Agric Food Chem. 2020 Jul 22;68(29):7710-7717. doi: 10.1021/acs.jafc.0c00294. Epub 2020 Jul 7. J Agric Food Chem. 2020. PMID: 32580548
-
De Novo Synthesis of Resveratrol from Sucrose by Metabolically Engineered Yarrowia lipolytica.Biomolecules. 2024 Jun 16;14(6):712. doi: 10.3390/biom14060712. Biomolecules. 2024. PMID: 38927115 Free PMC article.
-
De Novo Biosynthesis of Lutein in Yarrowia lipolytica.J Agric Food Chem. 2024 Mar 13;72(10):5348-5357. doi: 10.1021/acs.jafc.3c09080. Epub 2024 Feb 27. J Agric Food Chem. 2024. PMID: 38412053
-
Advanced Strategies for the Synthesis of Terpenoids in Yarrowia lipolytica.J Agric Food Chem. 2021 Mar 3;69(8):2367-2381. doi: 10.1021/acs.jafc.1c00350. Epub 2021 Feb 17. J Agric Food Chem. 2021. PMID: 33595318 Review.
-
[Advances in efficient biosynthesis of erythritol by metabolic engineering of Yarrowia lipolytica].Sheng Wu Gong Cheng Xue Bao. 2024 Mar 25;40(3):665-686. doi: 10.13345/j.cjb.230391. Sheng Wu Gong Cheng Xue Bao. 2024. PMID: 38545970 Review. Chinese.
References
-
- Qureshi A.A., Khan D.A., Mushtaq S., Ye S.Q., Xiong M., Qureshi N. delta-Tocotrienol feeding modulates gene expression of EIF2, mTOR, protein ubiquitination through multiple-signaling pathways in chronic hepatitis C patients. Lipids Health Dis. 2018;17:167. doi: 10.1186/s12944-018-0804-7. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
