

Combined Deletion of ZFP36L1 and ZFP36L2 Drives Superior Cytokine Production in T Cells at the Cost of Cell Fitness
- PMID: 40249077
- PMCID: PMC12007392
- DOI: 10.1002/eji.202451641
Combined Deletion of ZFP36L1 and ZFP36L2 Drives Superior Cytokine Production in T Cells at the Cost of Cell Fitness
Abstract
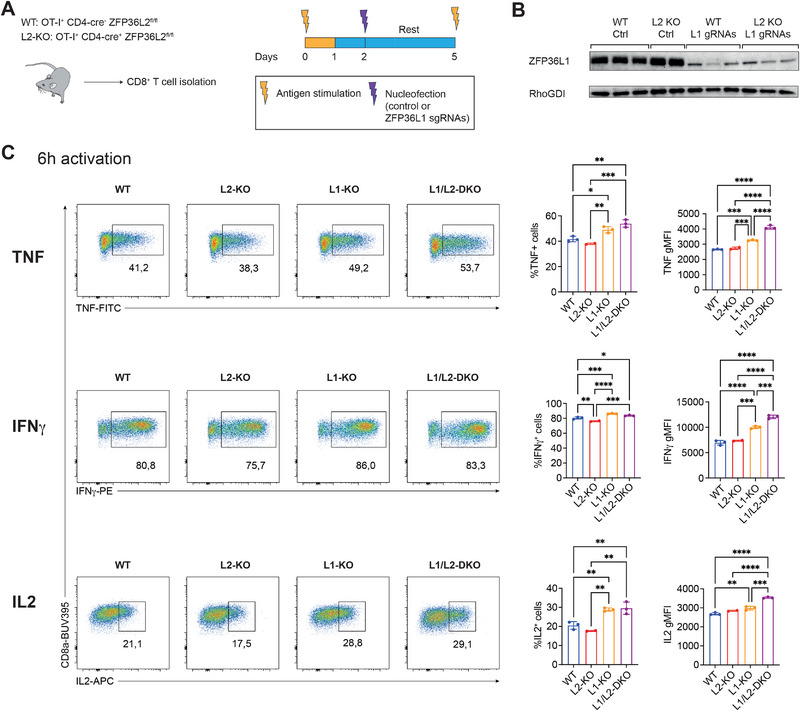
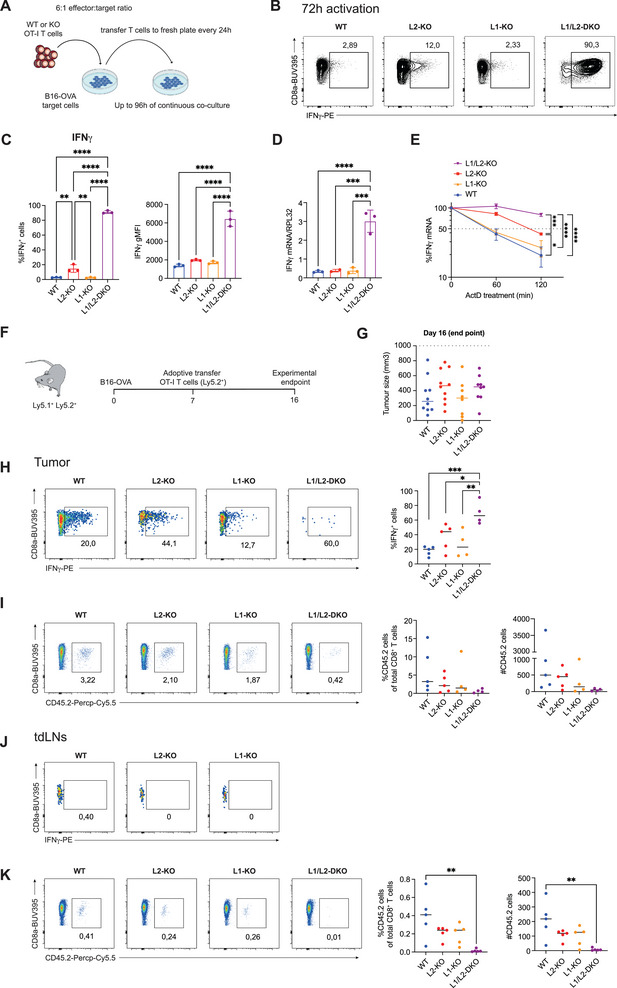
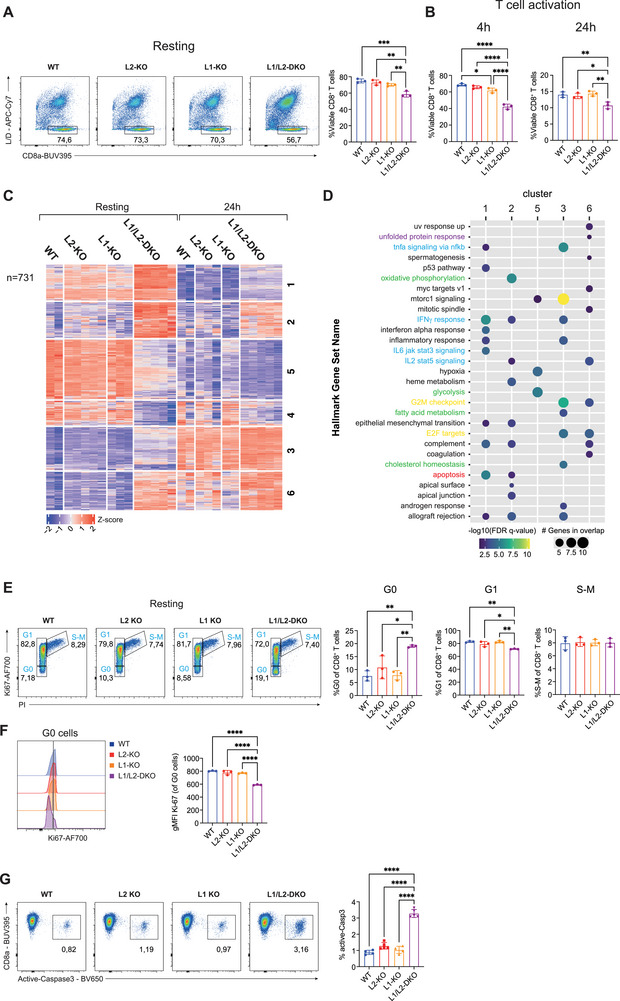
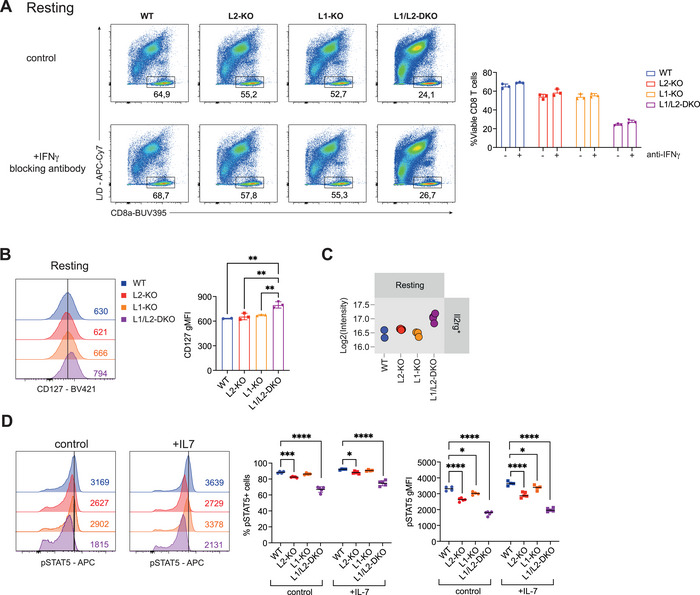
A key feature of cytotoxic CD8+ T cells for eliminating pathogens and malignant cells is their capacity to produce proinflammatory cytokines, which include TNF and IFNγ. Provided that these cytokines are highly toxic, a tight control of their production is imperative. RNA-binding proteins (RBPs) are essential for the fine-tuning of cytokine production. The role of the RBP ZFP36L1 and its sister protein ZFP36L2 herein has been established, but their relative contribution to cytokine production is not well understood. We here compared the effect of ZFP36L1 and ZFP36L2 single and double deficiency in murine effector CD8+ T cells. Whereas single deficient T cells significantly increased cytokine production, double deficiency completely unleashed the cytokine production. Not only the TNF production was substantially prolonged in double-deficient T cells. Also, the production of IFNγ reached unprecedented levels with >90% IFNγ-producing T cells compared with 3% in WT T cells after 3 days of continuous activation. This continuous cytokine production by double-deficient T cells was also observed in tumor-infiltrating lymphocytes in vivo, however, with no effect on tumor growth. ZFP36L1 and ZFP36L2 double deficiency resulted in decreased cell viability, impaired STAT5 signaling, and dysregulated cell cycle progression. In conclusion, while combined deletion in ZFP36L1 and ZFP36L2 can drive continuous cytokine production even upon chronic activation, safeguards are in place to counteract such super-cytokine producers.
© 2025 The Author(s). European Journal of Immunology published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Regulation of IFN-γ production by ZFP36L2 in T cells is time-dependent.Eur J Immunol. 2024 Oct;54(10):e2451018. doi: 10.1002/eji.202451018. Epub 2024 Jul 9. Eur J Immunol. 2024. PMID: 38980256
-
ZFP36-family RNA-binding proteins in regulatory T cells reinforce immune homeostasis.Nat Commun. 2025 May 6;16(1):4192. doi: 10.1038/s41467-025-58993-y. Nat Commun. 2025. PMID: 40328742 Free PMC article.
-
Deletion of the RNA-binding proteins ZFP36L1 and ZFP36L2 leads to perturbed thymic development and T lymphoblastic leukemia.Nat Immunol. 2010 Aug;11(8):717-24. doi: 10.1038/ni.1901. Epub 2010 Jul 11. Nat Immunol. 2010. PMID: 20622884 Free PMC article.
-
The ZFP36 family of RNA binding proteins regulates homeostatic and autoreactive T cell responses.Sci Immunol. 2022 Oct 28;7(76):eabo0981. doi: 10.1126/sciimmunol.abo0981. Epub 2022 Oct 21. Sci Immunol. 2022. PMID: 36269839 Free PMC article.
-
Time-dependent regulation of cytokine production by RNA binding proteins defines T cell effector function.Cell Rep. 2023 May 30;42(5):112419. doi: 10.1016/j.celrep.2023.112419. Epub 2023 Apr 18. Cell Rep. 2023. PMID: 37074914 Free PMC article.
References
-
- Ciuffreda D., Comte D., Cavassini M., et al., “Polyfunctional HCV‐specific T‐cell Responses Are Associated with Effective Control of HCV Replication,” European Journal of Immunology 38 (2008): 2665–2677. - PubMed
-
- Grivennikov S. I., Tumanov A. V., Liepinsh D. J., et al., “Distinct and Nonredundant in Vivo Functions of TNF Produced by T Cells and Macrophages/Neutrophils,” Immunity 22 (2005): 93–104. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
