

Cell lineage-resolved embryonic morphological map reveals signaling associated with cell fate and size asymmetry
- PMID: 40251161
- PMCID: PMC12008310
- DOI: 10.1038/s41467-025-58878-0
Cell lineage-resolved embryonic morphological map reveals signaling associated with cell fate and size asymmetry
Abstract
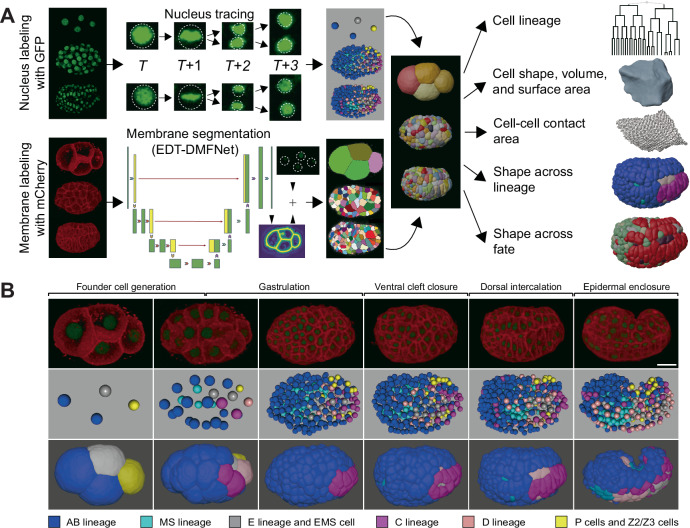
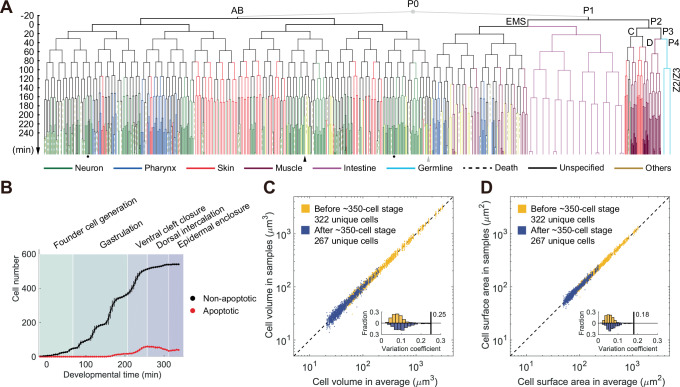
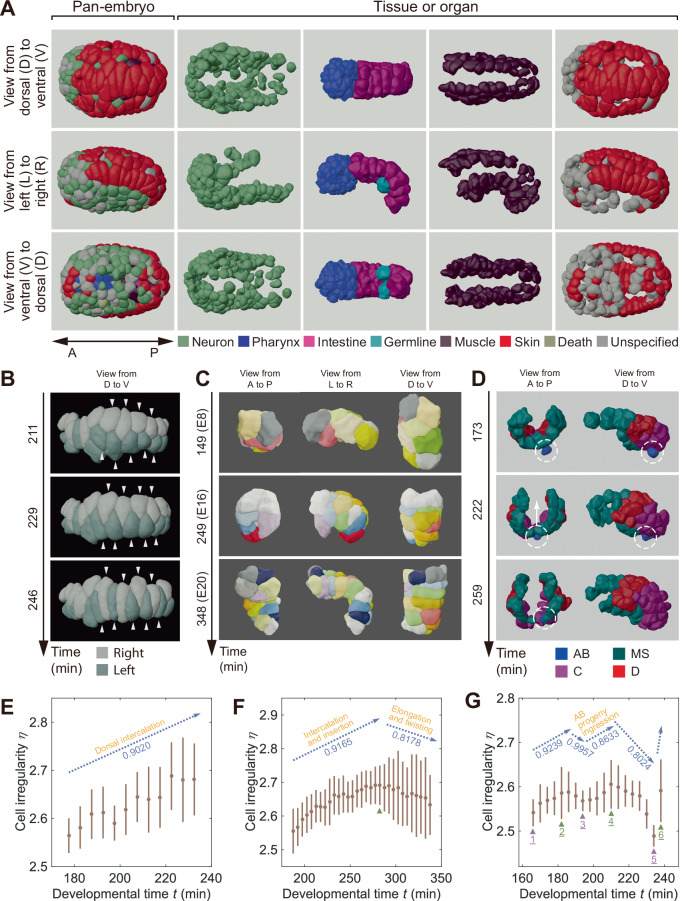
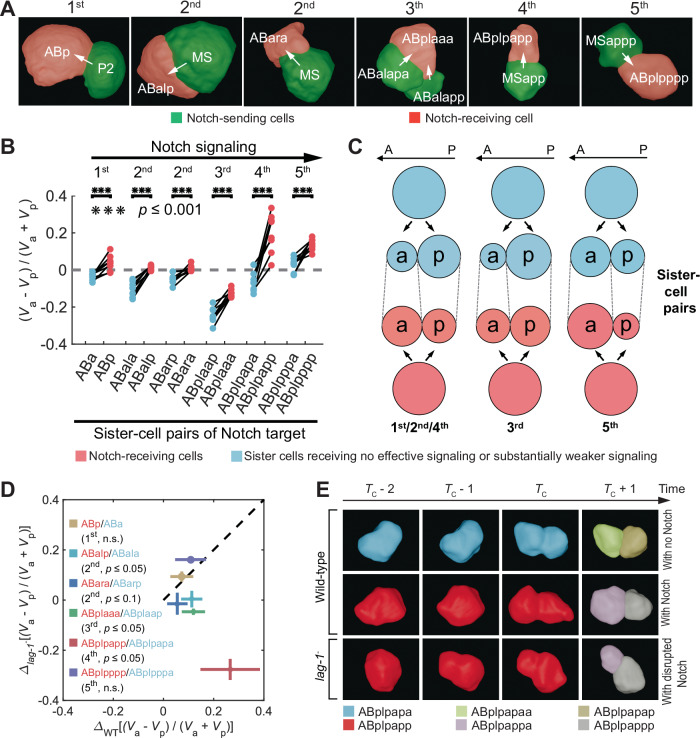
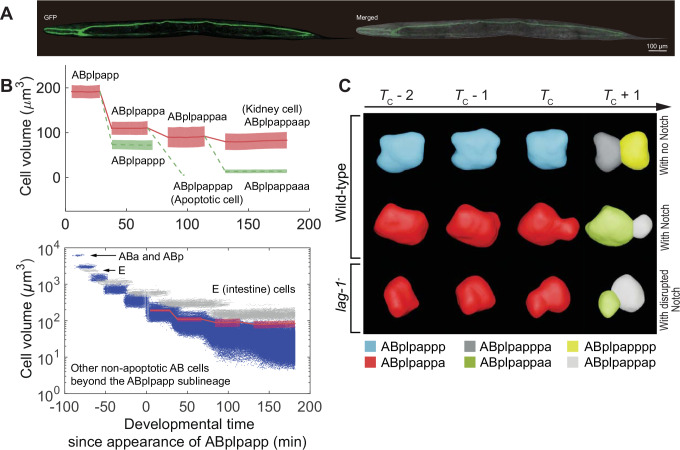
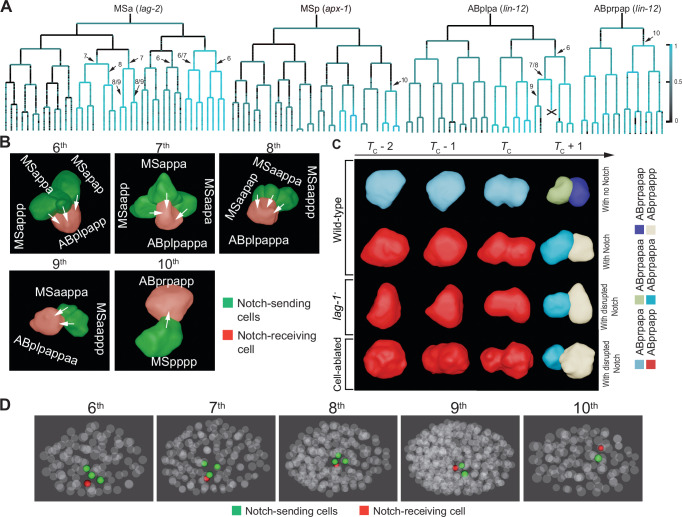
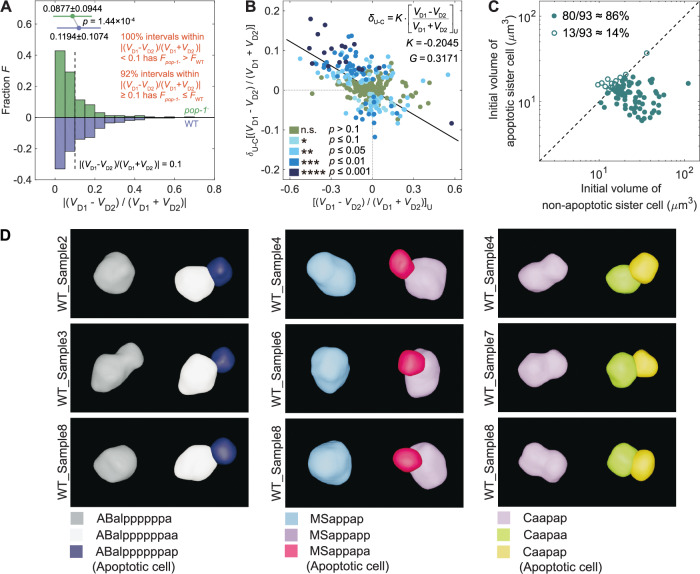
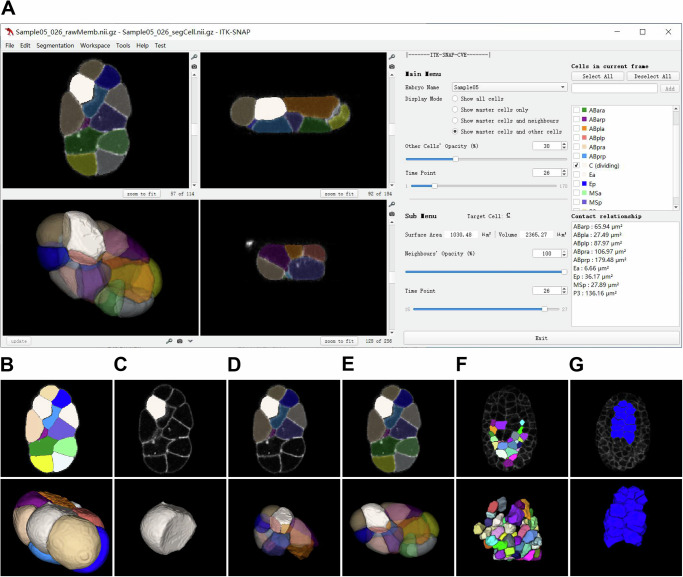
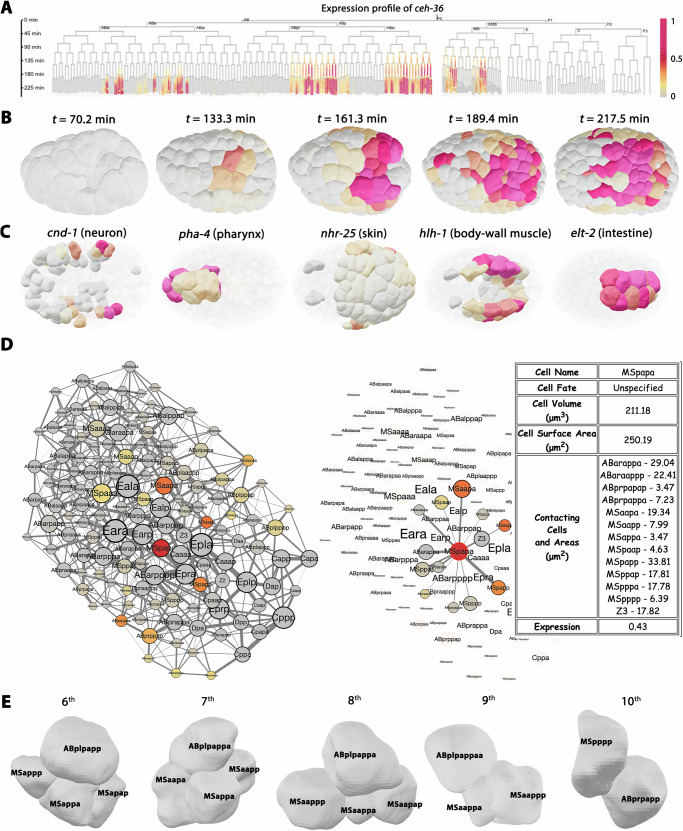
How cells change shape is crucial for the development of tissues, organs and embryos. However, studying these shape changes in detail is challenging. Here we present a comprehensive real-time cellular map that covers over 95% of the cells formed during Caenorhabditis elegans embryogenesis, featuring nearly 400,000 3D cell regions. This map includes information on each cell's identity, lineage, fate, shape, volume, surface area, contact area, and gene expression profiles, all accessible through our user-friendly software and website. Our map allows for detailed analysis of key developmental processes, including dorsal intercalation, intestinal formation, and muscle assembly. We show how Notch and Wnt signaling pathways, along with mechanical forces from cell interactions, regulate cell fate decisions and size asymmetries. Our findings suggest that repeated Notch signaling drives size disparities in the large excretory cell, which functions like a kidney. This work sets the stage for in-depth studies of the mechanisms controlling cell fate differentiation and morphogenesis.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
