

A host-pathogen metabolic synchrony that facilitates disease tolerance
- PMID: 40253414
- PMCID: PMC12009439
- DOI: 10.1038/s41467-025-59134-1
A host-pathogen metabolic synchrony that facilitates disease tolerance
Abstract
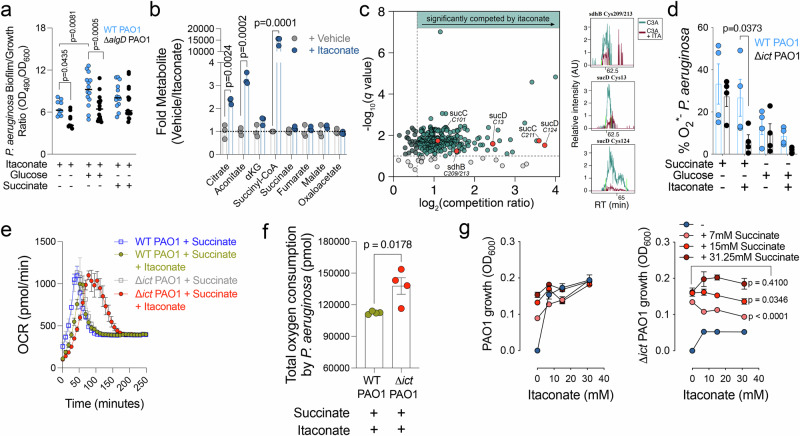
Disease tolerance mitigates organ damage from non-resolving inflammation during persistent infections, yet its underlying mechanisms remain unclear. Here we show, in a Pseudomonas aeruginosa pneumonia mouse model, that disease tolerance depends on the mitochondrial metabolite itaconate, which mediates cooperative host-pathogen interactions. In P. aeruginosa, itaconate modifies key cysteine residues in TCA cycle enzymes critical for succinate metabolism, inducing bioenergetic stress and promoting the formation biofilms that are less immunostimulatory and allow the bacteria to integrate into the local microbiome. Itaconate incorporates into the central metabolism of the biofilm, driving exopolysaccharide production-particularly alginate-which amplifies airway itaconate signaling. This itaconate-alginate interplay limits host immunopathology by enabling pulmonary glutamine assimilation, activating glutaminolysis, and thereby restrain detrimental inflammation caused by the inflammasome. Clinical sample analysis reveals that P. aeruginosa adapts to this metabolic environment through compensatory mutations in the anti-sigma-factor mucA, which restore the succinate-driven bioenergetics and disrupt the metabolic synchrony essential for sustaining disease tolerance.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures







References
MeSH terms
Substances
Grants and funding
- GT007268/Columbia University
- 1R21AI181780/Division of Intramural Research, National Institute of Allergy and Infectious Diseases (Division of Intramural Research of the NIAID)
- R35 GM146776/GM/NIGMS NIH HHS/United States
- R21 AI181780/AI/NIAID NIH HHS/United States
- RIQUEL21I0/Cystic Fibrosis Foundation (CF Foundation)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
