

Mannose and PMI depletion overcomes radiation resistance in HPV-negative head and neck cancer
- PMID: 40259370
- PMCID: PMC12013184
- DOI: 10.1186/s12964-025-02204-0
Mannose and PMI depletion overcomes radiation resistance in HPV-negative head and neck cancer
Abstract
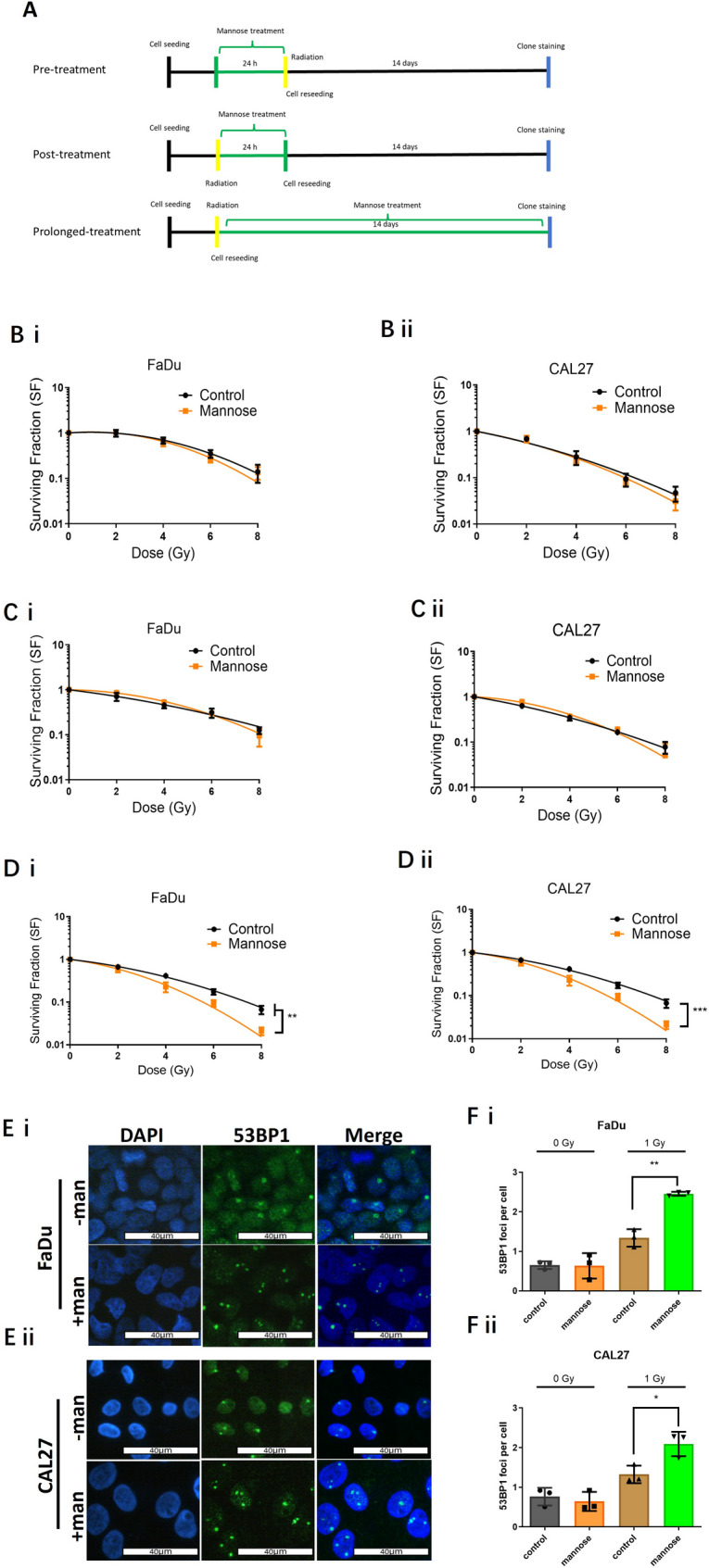
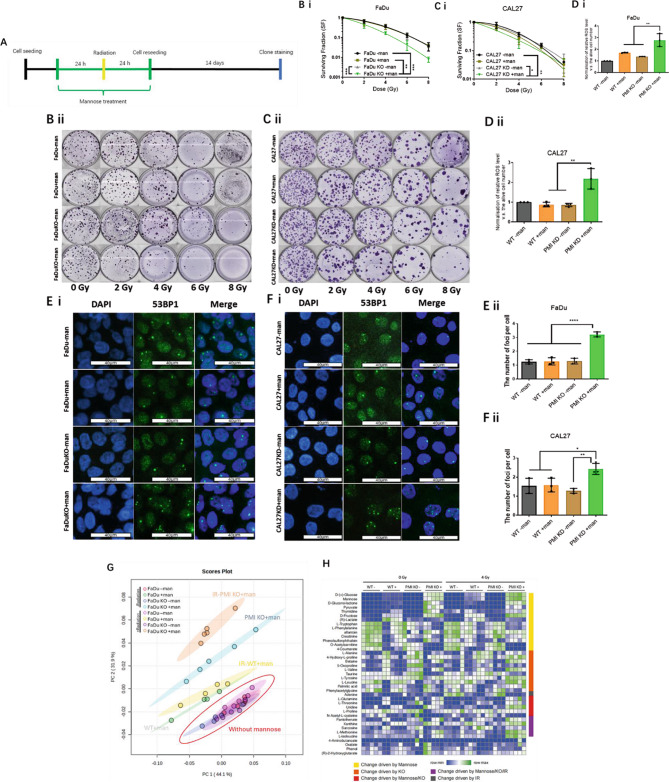
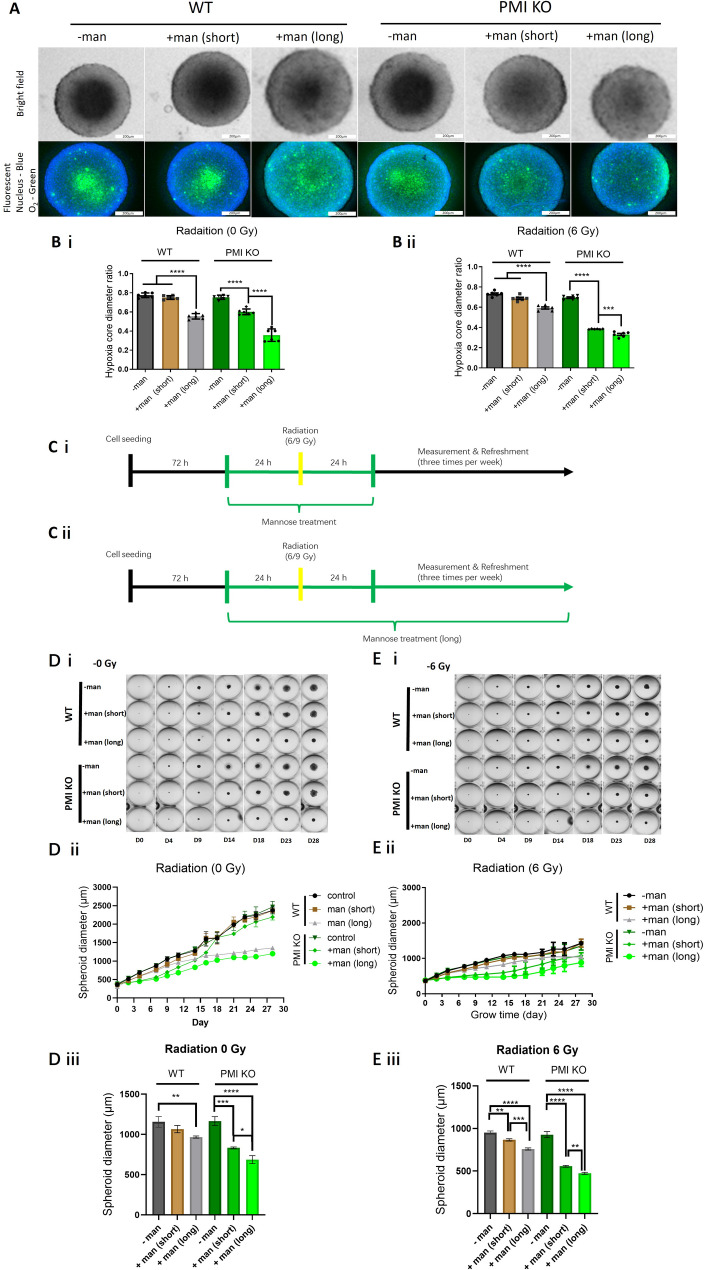
Radiotherapy is critical component of multidisciplinary cancer care, used as a primary and adjuvant treatment for patients with head and neck squamous cell carcinoma. This study investigates how mannose, a naturally occurring monosaccharide, combined with phosphomannose isomerase (PMI) depletion, enhances the sensitivity of HPV-negative head and neck tumour models to radiation. Isogenic PMI knockout models were generated by CRISPR/Cas9 gene editing, yielding a 20-fold increase in sensitivity to mannose in vitro, and causing significant tumour growth delay in vivo. This effect is driven by metabolic reprogramming, resulting in potent glycolytic suppression coupled with consistent depletion of ATP and glycolytic intermediates in PMI-depleted models. Functionally, these changes impede DNA damage repair following radiation, resulting in a significant increase in radiation sensitivity. Mannose and PMI ablation supressed both oxygen consumption rate and extracellular acidification, pushing cells towards a state of metabolic quiescence, effects contributing to increased radiation sensitivity under both normoxic and hypoxic conditions. In 3D-tumoursphere models, metabolic suppression by mannose and PMI depletion was shown to elevate intra-tumoursphere oxygen levels, contributing to significant in vitro oxygen-mediated radiosensitisation. These findings position PMI as a promising anti-tumour target, highlighting the potential of mannose as a metabolic radiosensitiser enhancing cancer treatment efficacy.
Keywords: Head and neck cancer; Mannose; Phosphomannose isomerase; Radiotherapy; Tumour metabolism.
Plain language summary
Radiotherapy is a vital treatment for head and neck cancer, however, many of these tumours poses areas that are poorly oxygenated, a feature that that causes resistance to radiotherapy. This study explores how mannose, a natural sugar, combined with reducing the activity of a protein called phosphomannose isomerase (PMI), makes HPV-negative head and neck tumours more sensitive to the effects of radiation. By dampening the ability of PMI to metabolise mannose, we report that the accumulation mannose metabolites disrupt tumour metabolism, depleting the tumour cell supply of energy (ATP), supressing the ability to repair DNA damage following radiation, and importantly helping to overcome resistance to radiation treatment caused by low-oxygen conditions. In 3D-tumour models, this combination resulted in increased tumoursphere oxygenation, further enhancing the effectiveness of radiation. These findings suggest PMI coupled with mannose as a promising target to improve cancer treatment outcomes.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures




References
-
- Rosenthal DI, Mohamed ASR, Garden AS, Morrison WH, El-Naggar AK, Kamal M, et al. Final report of a prospective randomized trial to evaluate the Dose-Response relationship for postoperative radiation therapy and pathologic risk groups in patients with head and neck cancer. Int J Radiat Oncol Biol Phys. 2017;98:1002–11. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
