

Challenges and advances for huntingtin detection in cerebrospinal fluid: in support of relative quantification
- PMID: 40259428
- PMCID: PMC12012996
- DOI: 10.1186/s40364-025-00772-4
Challenges and advances for huntingtin detection in cerebrospinal fluid: in support of relative quantification
Abstract
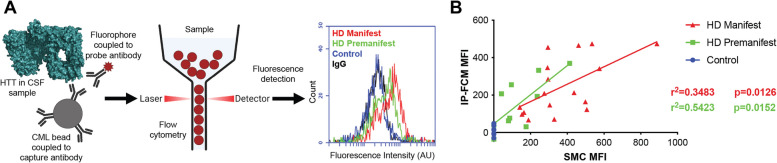
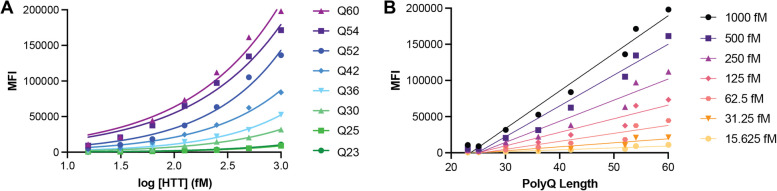
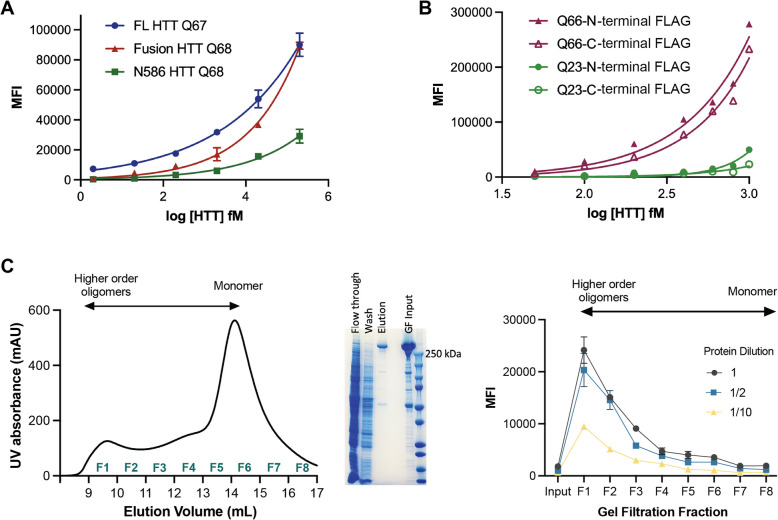
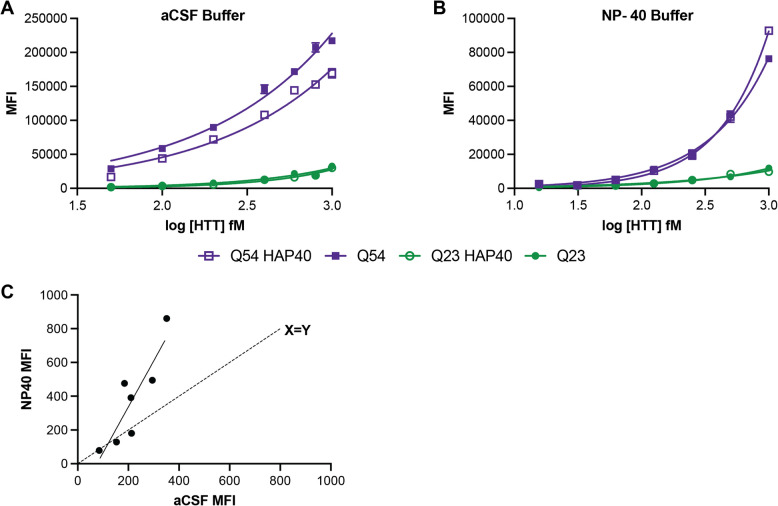
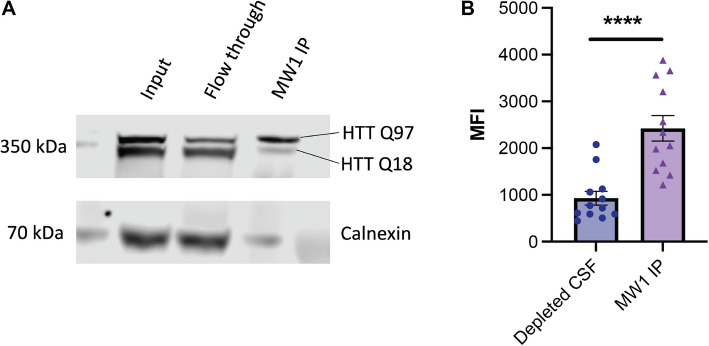
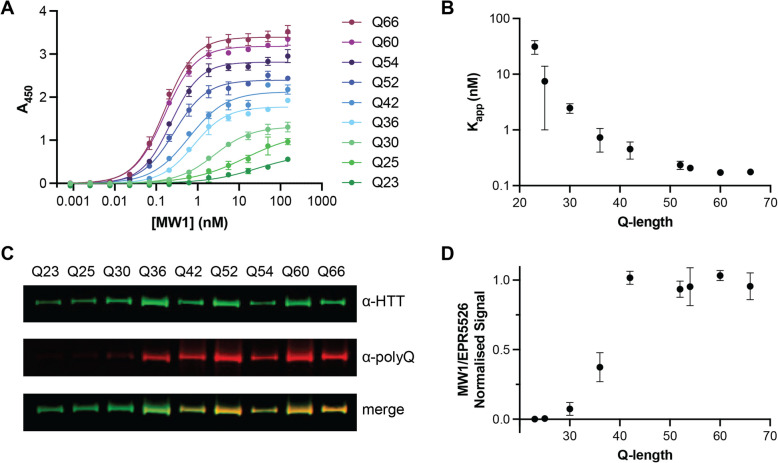
Huntington disease (HD) is a progressive and devastating neurodegenerative disease caused by expansion of a glutamine-coding CAG tract in the huntingtin (HTT) gene above a critical threshold of ~ 35 repeats resulting in expression of mutant HTT (mHTT). A promising treatment approach being tested in clinical trials is HTT lowering, which aims to reduce levels of the mHTT protein. Target engagement of these therapies in the brain are inferred using antibody-based assays that measure mHTT levels in the cerebrospinal fluid (CSF). These levels are typically reported as the absolute concentration of mHTT concentration, derived from a standard curve generated using a single protein standard. However, patient biofluids are a complex milieu containing different mHTT protein species, suggesting that absolute quantitation is challenging. As a result, a single recombinant protein standard may not be sufficient to interpret assay signal as molar mHTT concentration. In this study, we used immunoprecipitation and flow cytometry (IP-FCM) to investigate different factors that influence mHTT detection assay signal. Our results show that HTT protein fragmentation, protein-protein interactions, affinity tag positioning, oligomerization and polyglutamine tract length affect assay signal intensity. These findings indicate that absolute HTT quantitation in heterogeneous biological samples is not possible with current technologies using a single standard protein. We also explore the binding specificity of the MW1 anti-polyglutamine antibody, commonly used in these assays as a mHTT-selective reagent and demonstrate that mHTT binding is preferred but not specific. Furthermore, we find that MW1 depletion of mHTT for quantitation of wildtype HTT is not only incomplete, leaving residual mHTT, but also non-specific, resulting in pull down of some wildtype HTT protein. Based on these observations, we recommend that mHTT detection assays report only relative mHTT quantitation using normalized arbitrary units of assay signal intensity, rather than molar concentrations, in the assessment of central nervous system HTT lowering in ongoing clinical and preclinical studies. Further, we recommend that MW1-depletion not be used as a method for quantifying wildtype HTT protein and that detergent be consistently added to samples during testing.
Keywords: Biomarkers; Huntingtin Detection; Huntingtin Lowering Therapies; Huntington’s Disease.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Experiments were performed with the approval of the University of Central Florida Institute Animal Care and Use Committee (202400016) and Institutional Review Board (00002651). Consent for publication: Not applicable. Competing interests: B.R.L. reports previous roles as a scientific consultant with Alnylam, sRNAlytics, Teva, Roche/Genentech, Takeda, Triplet, Ionis, Novartis, Spark, Sage, Sintetica, LifeEdit, Design, Remix Therapeutics, UniQure, and PTC Therapeutics. B.R.L. is a co-founder and the CEO of Incisive Genetics Inc. B.R.L. is a founding Co-Editor-in-Chief of the Journal of Huntington’s Disease. D.R.L. is a paid biostatistical consultant to the Huntington Study Group, Sage Pharmaceuticals, Prilenia, Guidepoint Consultants, and Uniqure for designing Huntington’s Disease trials. M.R.H. serves on the boards of Ionis Pharmaceuticals, AbCellera and 89Bio.
Figures
Update of
-
Challenges and advances for huntingtin detection in cerebrospinal fluid: in support of relative quantification.bioRxiv [Preprint]. 2024 Sep 27:2024.09.25.614766. doi: 10.1101/2024.09.25.614766. bioRxiv. 2024. Update in: Biomark Res. 2025 Apr 21;13(1):63. doi: 10.1186/s40364-025-00772-4. PMID: 39386513 Free PMC article. Updated. Preprint.
References
-
- Benn CL, Sun T, Sadri-Vakili G, McFarland KN, DiRocco DP, Yohrling GJ, Clark TW, Bouzou B, Cha J-HJ. Huntingtin modulates transcription, occupies gene promoters in vivo, and binds directly to DNA in a polyglutamine-dependent manner. J Neurosci. 2008;28:10720–33. 10.1523/JNEUROSCI.2126-08.2008. - PMC - PubMed
-
- Bhattacharyya A, Trotta CR, Narasimhan J, Wiedinger KJ, Li W, Effenberger KA, Woll MG, Jani MB, Risher N, Yeh S, Cheng Y, Sydorenko N, Moon Y-C, Karp GM, Weetall M, Dakka A, Gabbeta V, Naryshkin NA, Graci JD, Tripodi T, Southwell A, Hayden M, Colacino JM, Peltz SW. Small molecule splicing modifiers with systemic HTT-lowering activity. Nat Commun. 2021;12:7299. 10.1038/s41467-021-27157-z. - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
