

FOXM1 cooperates with ERα to regulate functional β-cell mass
- PMID: 40261794
- PMCID: PMC12145799
- DOI: 10.1152/ajpendo.00438.2024
FOXM1 cooperates with ERα to regulate functional β-cell mass
Abstract
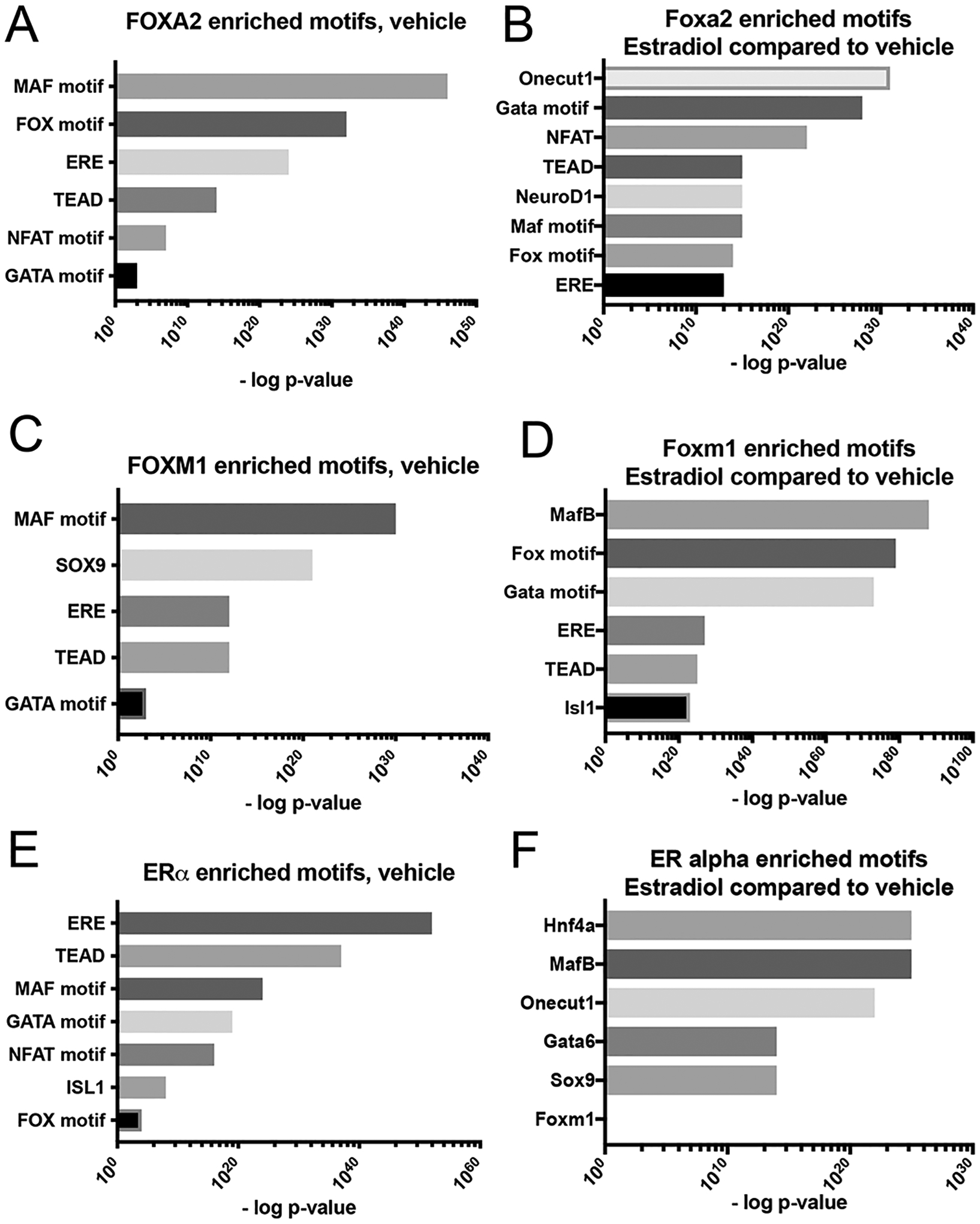
The transcription factor forkhead box (FOX)M1 regulates β-cell proliferation and insulin secretion. Our previous work demonstrates that expressing a constitutively active form of FOXM1 (FOXM1*) in β-cells increases β-cell function, proliferation, and mass in male mice. However, in contrast to what is observed in males, we demonstrate here that in female mice expression of FOXM1* in β-cells does not affect β-cell proliferation or glucose tolerance. Similarly, FOXM1* transduction of male but not female human islets enhances insulin secretion in response to elevated glucose. We therefore examined the mechanism behind this sexual dimorphism. Estrogen contributes to diabetes susceptibility differences between males and females, and estrogen receptor (ER)α is the primary mediator of β-cell estrogen signaling. Moreover, in breast cancer cells, ERα and FOXM1 work together to drive gene expression. We therefore examined whether FOXM1 and ERα functionally interact in β-cells. FOXM1* rescued elevated fasting glucose, glucose intolerance, and homeostatic model assessment of β-cell function (HOMA-B) in female mice with a β-cell-specific ERα deletion. Furthermore, in the presence of estrogen, the FOXM1 and ERα cistromes exhibit significant overlap in βTC6 β-cells. In addition, FOXM1 and ERα binding sites frequently occur in complex enhancers co-occupied by other islet transcription factors. These data indicate that FOXM1 and nuclear ERα cooperate to regulate β-cell function and suggest a general mechanism contributing to the lower incidence of diabetes observed in women.NEW & NOTEWORTHY Here we investigate why the effects of increasing FOXM1 activity in β-cells observed in male mice are not seen in female mice. ERα likely collaborates with FOXM1 and other transcription factors to enhance gene expression related to β-cell function. Higher estrogen levels in females may contribute to their increased insulin secretion and the more severe consequences of losing transcription factors like FOXM1 in males. Overall, these findings shed light on sex differences in diabetes susceptibility.
Keywords: Foxm1; beta cells; estrogen receptor.
Conflict of interest statement
Declaration of Interest
The authors declare no conflicts of interest.
Figures









References
-
- Saeedi P, Petersohn I, Salpea P, Malanda B, Karuranga S, Unwin N, Colagiuri S, Guariguata L, Motala AA, Ogurtsova K, Shaw JE, Bright D, Williams R, and Committee IDFDA. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res Clin Pract 157: 107843, 2019. - PubMed
-
- Kim JH, and Saxton AM. The TALLYHO mouse as a model of human type 2 diabetes. Methods Mol Biol 933: 75–87, 2012. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous