

HU promotes higher order chromosome organization and influences DNA replication rates in Streptococcus pneumoniae
- PMID: 40263708
- PMCID: PMC12014288
- DOI: 10.1093/nar/gkaf312
HU promotes higher order chromosome organization and influences DNA replication rates in Streptococcus pneumoniae
Abstract
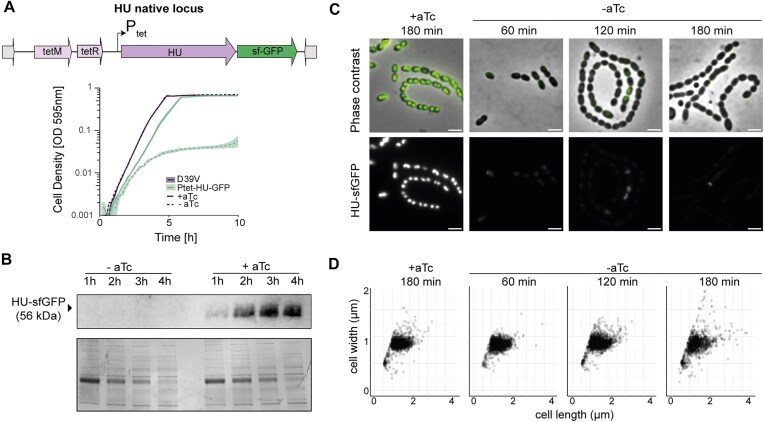
Nucleoid-associated proteins (NAPs) are crucial for maintaining chromosomal compaction and architecture, and are actively involved in DNA replication, recombination, repair, and gene regulation. In Streptococcus pneumoniae, the role of the highly conserved NAP HU in chromosome conformation has not yet been investigated. Here, we use a multi-scale approach to explore HU's role in chromosome conformation and segregation dynamics. By combining superresolution microscopy and whole-genome-binding analysis, we describe the nucleoid as a dynamic structure where HU binds transiently across the entire nucleoid, with a preference for the origin of replication over the terminus. Reducing cellular HU levels impacts nucleoid maintenance and disrupts nucleoid scaling with cell size, similar to the distortion caused by fluoroquinolones, supporting its requirement for maintaining DNA supercoiling. Furthermore, in cells lacking HU, the replication machinery is misplaced, preventing cells from initiating and proceeding with ongoing replication. Chromosome conformation capture coupled to deep sequencing (Hi-C) revealed that HU is required to maintain cohesion between the two chromosomal arms, similar to the structural maintenance of chromosome complex. Together, we show that by promoting long-range chromosome interactions and supporting the architecture of the domain encompassing the origin, HU is essential for chromosome integrity and the intimately related processes of replication and segregation.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
None declared.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
