

Cerebellar Purkinje cells control posture in larval zebrafish (Danio rerio)
- PMID: 40272244
- PMCID: PMC12021414
- DOI: 10.7554/eLife.97614
Cerebellar Purkinje cells control posture in larval zebrafish (Danio rerio)
Abstract
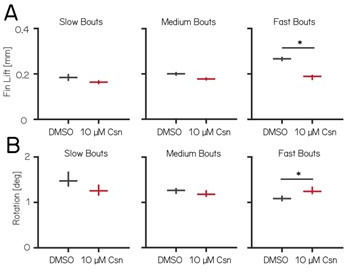
Cerebellar dysfunction leads to postural instability. Recent work in freely moving rodents has transformed investigations of cerebellar contributions to posture. However, the combined complexity of terrestrial locomotion and the rodent cerebellum motivate new approaches to perturb cerebellar function in simpler vertebrates. Here, we adapted a validated chemogenetic tool (TRPV1/capsaicin) to describe the role of Purkinje cells - the output neurons of the cerebellar cortex - as larval zebrafish swam freely in depth. We achieved both bidirectional control (activation and ablation) of Purkinje cells while performing quantitative high-throughput assessment of posture and locomotion. Activation modified postural control in the pitch (nose-up/nose-down) axis. Similarly, ablations disrupted pitch-axis posture and fin-body coordination responsible for climbs. Postural disruption was more widespread in older larvae, offering a window into emergent roles for the developing cerebellum in the control of posture. Finally, we found that activity in Purkinje cells could individually and collectively encode tilt direction, a key feature of postural control neurons. Our findings delineate an expected role for the cerebellum in postural control and vestibular sensation in larval zebrafish, establishing the validity of TRPV1/capsaicin-mediated perturbations in a simple, genetically tractable vertebrate. Moreover, by comparing the contributions of Purkinje cell ablations to posture in time, we uncover signatures of emerging cerebellar control of posture across early development. This work takes a major step towards understanding an ancestral role of the cerebellum in regulating postural maturation.
Keywords: balance; imaging; neuroscience; postural control; purkinje cells; vestibular system; zebrafish.
© 2024, Auer et al.
Conflict of interest statement
FA, KN, KM, MH, DS No competing interests declared
Figures













Update of
-
Cerebellar Purkinje Cells Control Posture in Larval Zebrafish (Danio rerio).bioRxiv [Preprint]. 2024 Nov 9:2023.09.12.557469. doi: 10.1101/2023.09.12.557469. bioRxiv. 2024. Update in: Elife. 2025 Apr 24;13:RP97614. doi: 10.7554/eLife.97614. PMID: 37745506 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
