

Anti-Tumour Immunity Relies on Targeting Tissue Homeostasis Through Monocyte-Driven Responses Rather Than Direct Tumour Cytotoxicity
- PMID: 40272245
- PMCID: PMC12020664
- DOI: 10.1111/liv.70110
Anti-Tumour Immunity Relies on Targeting Tissue Homeostasis Through Monocyte-Driven Responses Rather Than Direct Tumour Cytotoxicity
Abstract
Background: Metabolic dysfunction-associated fatty liver disease (MAFLD) can progress to hepatocellular carcinoma (HCC), yet the immune mechanisms driving this transition remain unclear.
Methods: In a chronic Western diet (WD) mouse model, we performed single-nuclei RNA sequencing to track MAFLD progression into HCC and subsequent tumour inhibition upon dietary correction.
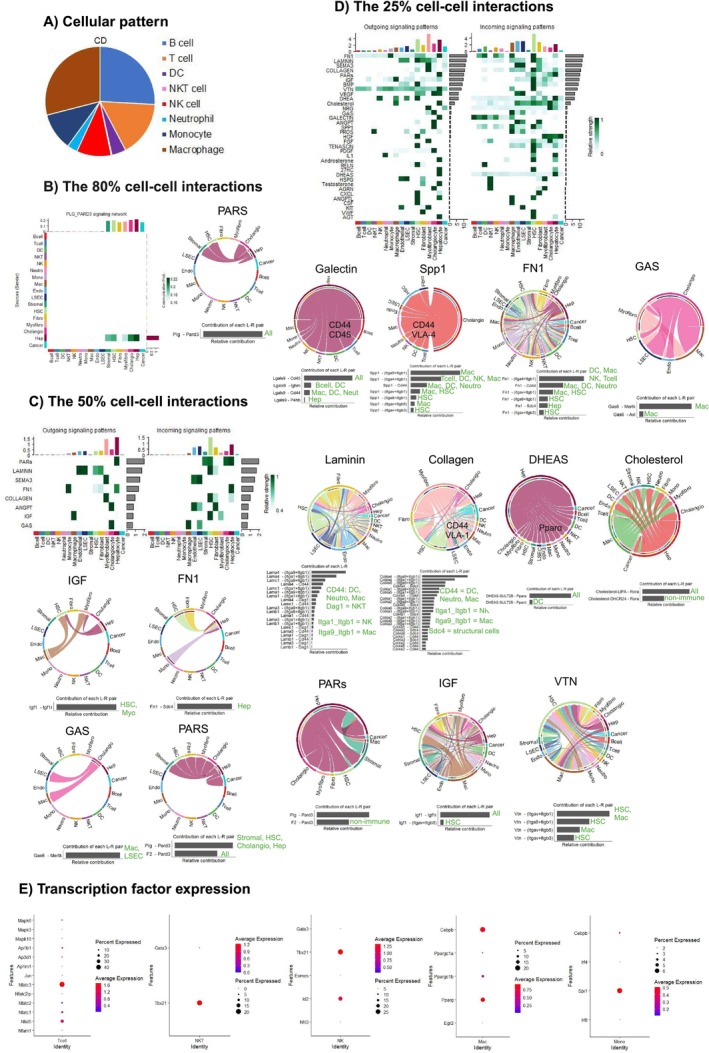
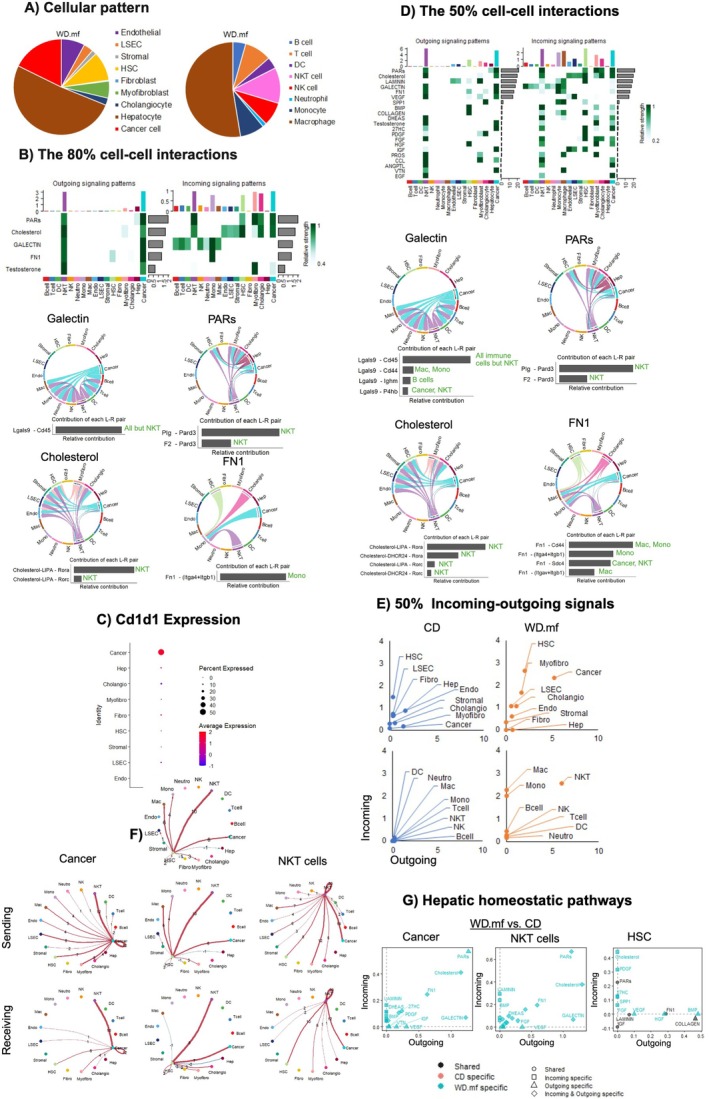
Results: Carcinogenesis begins during MAFLD, with tumour cells entering dormancy when HCC is mitigated. Rather than purely tolerogenic, the liver actively engages immune responses targeting myofibroblasts, fibroblasts and hepatocytes to maintain tissue homeostasis. Cytotoxic cells contribute to the turnover of liver cells but do not primarily target the tumour. NKT cells predominate under chronic WD, while monocytes join them in HCC progression on a WD. Upon dietary correction, monocyte-driven immunity confers protection against HCC through targeting tissue homeostatic pathways and antioxidant mechanisms. Crucially, liver tissue response-not merely immune activation-dictates whether tumours grow or regress, emphasising the importance of restoring liver tissue integrity. Also, protection against HCC is linked to a distinct immunological pattern, differing from healthy controls, underscoring the need for immune reprogramming.
Conclusion: These findings reveal the dual roles of similar pathways, where immune patterns targeting different cells shape distinct outcomes. Restoring tissue homeostasis and regeneration creates a tumour-hostile microenvironment, whereas tumour-directed approaches fail to remodel the TME. This underscores the need for tissue remodelling strategies in cancer prevention and treatment.
Keywords: cancer dormancy; hepatocellular carcinoma; inflammation; network medicine; nonalcoholic fatty liver disease; stromal cells; systems immunology.
© 2025 The Author(s). Liver International published by John Wiley & Sons Ltd.
Conflict of interest statement
A.J.S. and F.M. hold a patent on the DIAMOND mouse model, PCT/US2016/056506. All other authors do not have any competing interest.
Figures






Update of
-
Anti-tumor immunity relies on targeting tissue homeostasis through monocyte-driven responses rather than direct tumor cytotoxicity.bioRxiv [Preprint]. 2025 Mar 6:2024.06.12.598563. doi: 10.1101/2024.06.12.598563. bioRxiv. 2025. Update in: Liver Int. 2025 May;45(5):e70110. doi: 10.1111/liv.70110. PMID: 38903113 Free PMC article. Updated. Preprint.
Similar articles
-
Anti-tumor immunity relies on targeting tissue homeostasis through monocyte-driven responses rather than direct tumor cytotoxicity.bioRxiv [Preprint]. 2025 Mar 6:2024.06.12.598563. doi: 10.1101/2024.06.12.598563. bioRxiv. 2025. Update in: Liver Int. 2025 May;45(5):e70110. doi: 10.1111/liv.70110. PMID: 38903113 Free PMC article. Updated. Preprint.
-
Single-cell RNA sequencing reveals a reprogramming of hepatic immune cells and a protective role for B cells in MASH-driven HCC.Hepatol Commun. 2025 Apr 21;9(5):e0668. doi: 10.1097/HC9.0000000000000668. eCollection 2025 May 1. Hepatol Commun. 2025. PMID: 40257356 Free PMC article.
-
Metabolic reprogramming of immune cells: Shaping the tumor microenvironment in hepatocellular carcinoma.Cancer Med. 2021 Sep;10(18):6374-6383. doi: 10.1002/cam4.4177. Epub 2021 Aug 13. Cancer Med. 2021. PMID: 34390203 Free PMC article. Review.
-
Adaptive immunity suppresses formation and progression of diethylnitrosamine-induced liver cancer.Gut. 2012 Dec;61(12):1733-43. doi: 10.1136/gutjnl-2011-301116. Epub 2012 Jan 20. Gut. 2012. PMID: 22267597 Free PMC article.
-
Neutrophils: Driving inflammation during the development of hepatocellular carcinoma.Cancer Lett. 2021 Dec 1;522:22-31. doi: 10.1016/j.canlet.2021.09.011. Epub 2021 Sep 10. Cancer Lett. 2021. PMID: 34517084 Review.
Cited by
-
Bridging the Gap in Breast Cancer Dormancy: Models, Mechanisms, and Translational Challenges.Pharmaceuticals (Basel). 2025 Jun 26;18(7):961. doi: 10.3390/ph18070961. Pharmaceuticals (Basel). 2025. PMID: 40732251 Free PMC article. Review.
-
Tumor-associated bacteria activate PRDX1-driven glycolysis to promote immune evasion and PD-1 antibody resistance in hepatocellular carcinoma.Front Microbiol. 2025 Jul 7;16:1599691. doi: 10.3389/fmicb.2025.1599691. eCollection 2025. Front Microbiol. 2025. PMID: 40693141 Free PMC article.
-
Redefining the Immunobiology of Organ Transplantation for New Clinical Horizons.Scand J Immunol. 2025 Aug;102(2):e70045. doi: 10.1111/sji.70045. Scand J Immunol. 2025. PMID: 40717391 Free PMC article. Review.
References
-
- Tacke F., “Targeting Hepatic Macrophages to Treat Liver Diseases,” Journal of Hepatology 66 (2017): 1300–1312. - PubMed
-
- Nakashima H., Inui T., Habu Y., et al., “Activation of Mouse Natural Killer T Cells Accelerates Liver Regeneration After Partial Hepatectomy,” Gastroenterology 131 (2006): 1573–1583. - PubMed
-
- Anders R. A., Subudhi S. K., Wang J., Pfeffer K., and Fu Y.‐X., “Contribution of the Lymphotoxin Beta Receptor to Liver Regeneration,” Journal of Immunology 175 (2005): 1295–1300. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
