

Sex and sex hormonal regulation of the atrial inward rectifier potassium current (IK1): insights into potential pro-arrhythmic mechanisms
- PMID: 40272446
- PMCID: PMC12310280
- DOI: 10.1093/cvr/cvaf074
Sex and sex hormonal regulation of the atrial inward rectifier potassium current (IK1): insights into potential pro-arrhythmic mechanisms
Abstract
Aims: Pronounced sex-differences are known in the incidence of atrial fibrillation (AF). In this study, we aimed to investigate the atrial electrophysiological properties that may underlie sex-differences in AF incidence in the younger population, focusing on IK1, a cardiac ion current important for action potential (AP) stability and triggered activity.
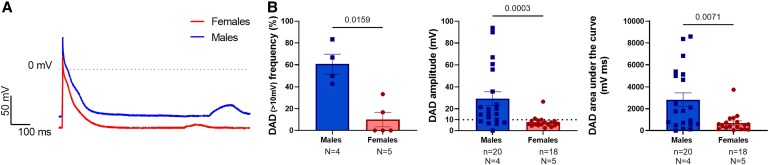
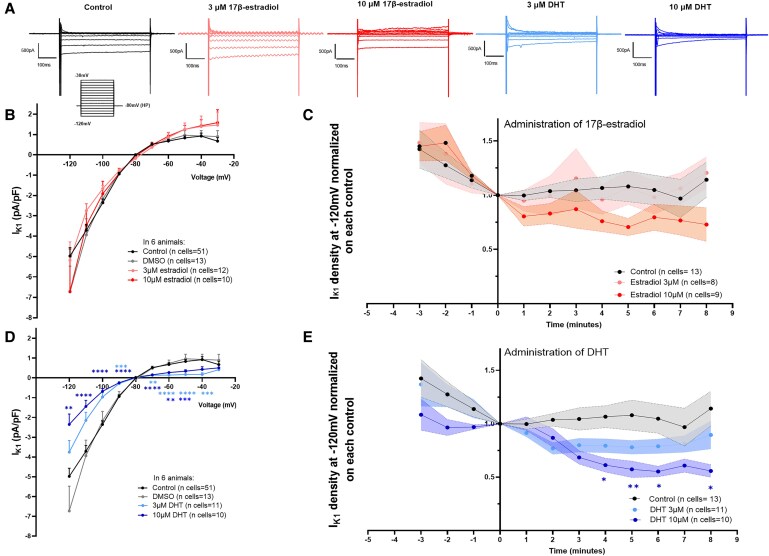
Methods and results: We assessed sex-differences in P-wave morphology in 12-lead ECG in healthy young New Zealand White rabbits. Males presented longer PWD and larger P-wave area compared to females. Patch-clamp experiments were performed in isolated rabbit atrial cardiomyocytes (CMs). Male atrial CMs presented higher delayed after depolarizations (DAD) incidence, amplitude, and area under the curve (AUC) than females, potentially facilitating the presence of atrial triggered activity in males. Male atrial CMs showed a less hyperpolarized resting membrane potential (RMP), a 50% smaller IK1, and a 26% reduction in Kir2.1 protein expression, a pore forming subunit of IK1, than females. Dihydrotestosterone (DHT) effects were investigated acutely and semi-chronically ex vivo. Experiments showed that the sex-difference in IK1 could be mimicked by DHT. In female atrial CMs, acute and semi-chronic (24 h) DHT administration reduced IK1. In the presence of a PKC-inhibitor, DHT-mediated IK1 reduction was not observed in atrial female CMs, suggesting it to be PKC-mediated. Chronic DHT-effects were investigated in vivo in female rabbits after hormone-releasing pellet implantation. After 2 weeks, animals showed a significantly prolonged and larger P-wave, a smaller atrial IK1 and a trend towards an increased DAD amplitude and AUC.
Conclusion: Sex impacts on atrial electrophysiology, leading to sex-differences in P-wave morphology, triggered activity, RMP, and IK1. These sex-differences can be mimicked by sex hormone-treatment, suggesting that sex hormones-particularly DHT-play a pivotal role in mediating sex-differences in atrial electrophysiology. Such sex-differences might impact on the propensity to develop AF, particularly in the younger population.
Keywords: Atrial fibrillation; IK1; Sex differences; Sex hormones.
© The Author(s) 2025. Published by Oxford University Press on behalf of the European Society of Cardiology.
Conflict of interest statement
Conflict of interest: U.S. received consultancy fees or honoraria from Università della Svizzera Italiana (USI, Switzerland) and Roche Diagnostics (Switzerland). U.S. received research grants from EP Solutions Inc. (Switzerland), Roche Diagnostics (Switzerland) and Bayer Healthcare (Germany). U.S. is co-founder and shareholder of YourRhythmics BV, a spin-off company of the University Maastricht.
Figures






Comment in
-
Testosterone and atrial fibrillation: does the dose make the poison?Cardiovasc Res. 2025 Jul 31;121(8):1144-1145. doi: 10.1093/cvr/cvaf113. Cardiovasc Res. 2025. PMID: 40579579 No abstract available.
References
-
- Andrade JG, Deyell MW, Lee AYK, Macle L. Sex differences in atrial fibrillation. Can J Cardiol 2018;34:429–436. - PubMed
-
- Benjamin EJ, Levy D, Vaziri SM, D'Agostino RB, Belanger AJ, Wolf PA. Independent risk factors for atrial fibrillation in a population-based cohort. The framingham heart study. JAMA 1994;271:840–844. - PubMed
-
- Friberg J, Scharling H, Gadsbøll N, Truelsen T, Jensen GB; Copenhagen City Heart Study . Comparison of the impact of atrial fibrillation on the risk of stroke and cardiovascular death in women versus men (The Copenhagen City Heart Study). Am J Cardiol 2004;94:889–894. - PubMed
-
- Odening KE, Deiß S, Dilling-Boer D, Didenko M, Eriksson U, Nedios S, Ng FS, Roca Luque I, Sanchez Borque P, Vernooy K, Wijnmaalen AP, Yorgun H. Mechanisms of sex differences in atrial fibrillation: role of hormones and differences in electrophysiology, structure, function, and remodelling. Europace 2019;21:366–376. - PubMed
