

Encoding and decoding selectivity and promiscuity in the human chemokine-GPCR interaction network
- PMID: 40273912
- PMCID: PMC12435897
- DOI: 10.1016/j.cell.2025.03.046
Encoding and decoding selectivity and promiscuity in the human chemokine-GPCR interaction network
Abstract
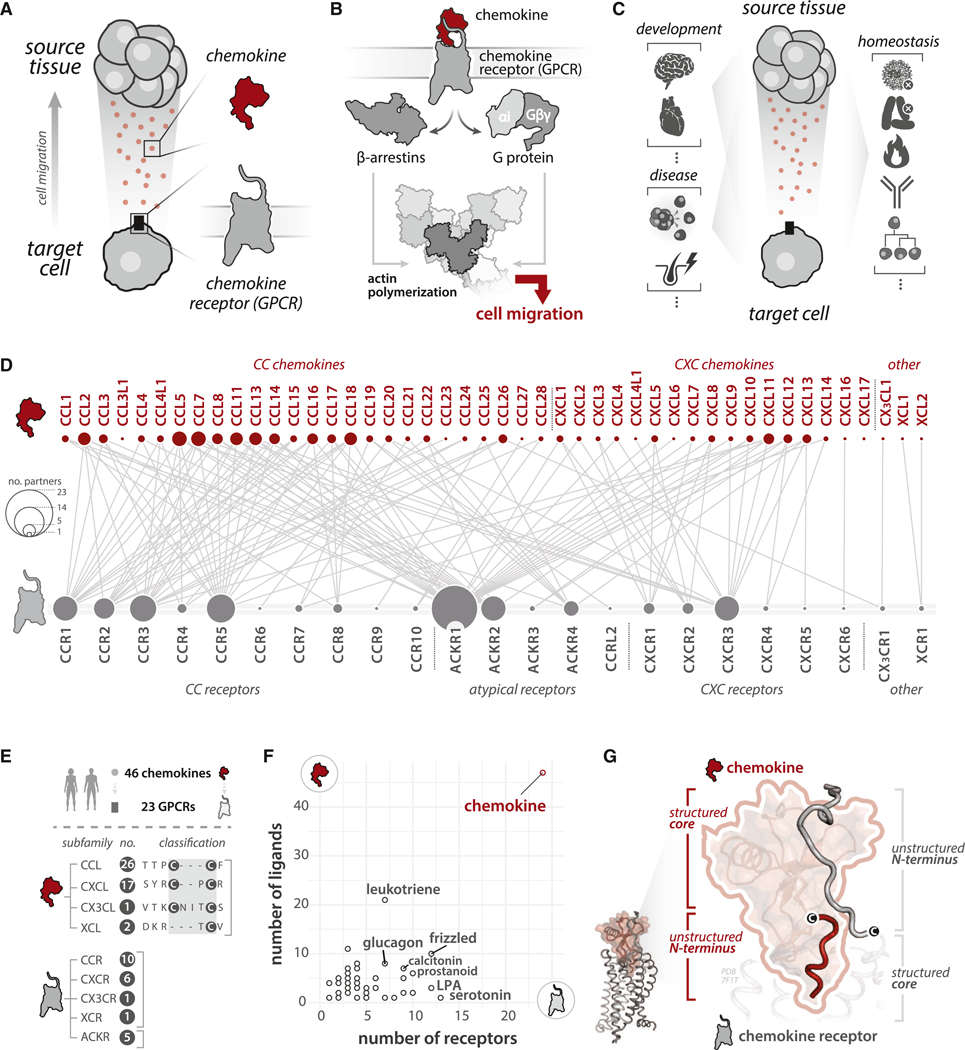
In humans, selective and promiscuous interactions between 46 secreted chemokine ligands and 23 cell surface chemokine receptors of the G-protein-coupled receptor (GPCR) family form a complex network to coordinate cell migration. While chemokines and their GPCRs each share common structural scaffolds, the molecular principles driving selectivity and promiscuity remain elusive. Here, we identify conserved, semi-conserved, and variable determinants (i.e., recognition elements) that are encoded and decoded by chemokines and their receptors to mediate interactions. Selectivity and promiscuity emerge from an ensemble of generalized ("public/conserved") and specific ("private/variable") determinants distributed among structured and unstructured protein regions, with ligands and receptors recognizing these determinants combinatorially. We employ these principles to engineer a viral chemokine with altered GPCR coupling preferences and provide a web resource to facilitate sequence-structure-function studies and protein design efforts for developing immuno-therapeutics and cell therapies.
Keywords: GPCR; chemokine; chemotaxis; data science; machine learning; polymorphism; protein-protein interaction; selectivity determinants; short linear motif; unstructured protein.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.F.V and F.C.P. have ownership interests in Protein Foundry, LLC and XLock Biosciences, Inc.
Figures






References
-
- Ma Q, Jones D, Borghesani PR, Segal RA, Nagasawa T, Kishimoto T, Bronson RT, and Springer TA (1998). Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice. Proc. Natl. Acad. Sci. USA 95, 9448–9453. 10.1073/pnas.95.16.9448. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
