

Scalable Production of Recombinant Adeno-Associated Virus Vectors Expressing Soluble Viral Receptors for Broad-Spectrum Inhibition of Porcine Reproductive and Respiratory Syndrome Virus Type 2
- PMID: 40284868
- PMCID: PMC12031001
- DOI: 10.3390/vetsci12040366
Scalable Production of Recombinant Adeno-Associated Virus Vectors Expressing Soluble Viral Receptors for Broad-Spectrum Inhibition of Porcine Reproductive and Respiratory Syndrome Virus Type 2
Abstract
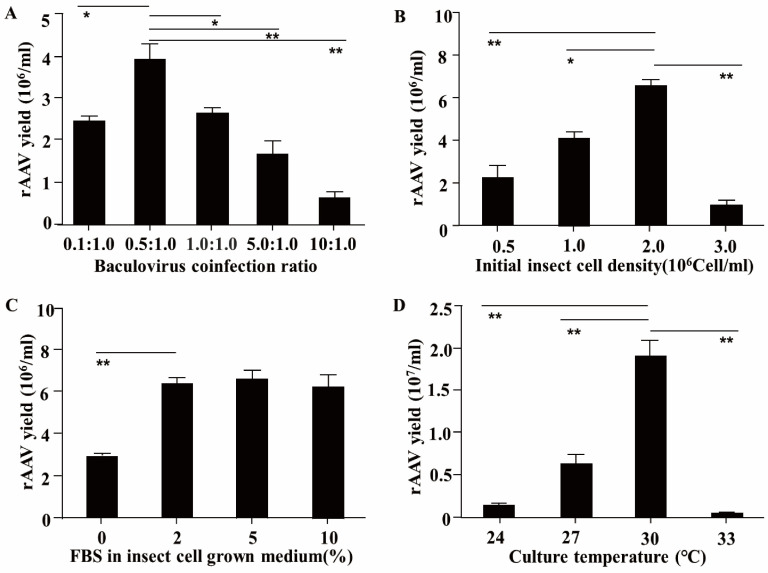
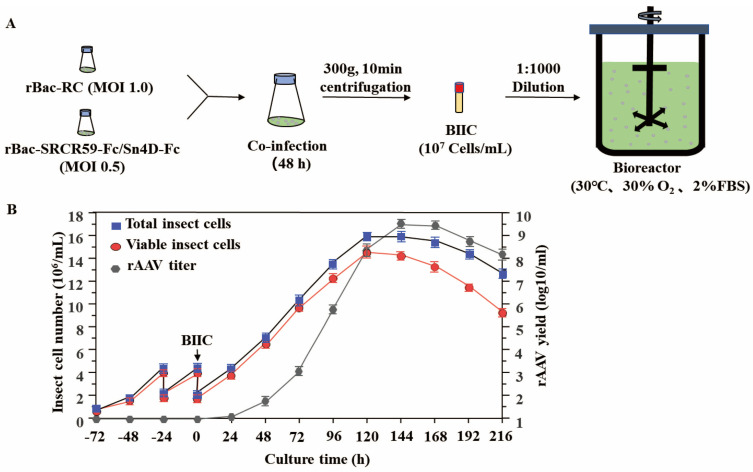
Porcine reproductive and respiratory syndrome virus (PRRSV) continues to be a major threat to the global swine industry, causing significant economic losses. To address this, we developed a scalable recombinant adeno-associated virus (rAAV)-based strategy for the delivery of soluble viral receptors (SVRs) to treat and potentially eliminate PRRSV infections. This strategy involves fusing the virus-binding domains of two key cellular receptors, sialoadhesin (Sn4D) and CD163 (SRCR5-9), with an Fc fragment. We then used an insect cell-baculovirus expression vector system to produce the rAAV-SRCR59-Fc/Sn4D-Fc vector. Through a series of optimizations, we determined the best conditions for rAAV production, including a baculovirus co-infection ratio of 0.5:1.0, an initial insect cell density of 2.0 × 106 cells/mL, a fetal bovine serum concentration of 2%, and a culture temperature of 30 °C. Under these optimized conditions, we achieved a high titer of rAAV-SRCR59-Fc/Sn4D-Fc in a 2 L bioreactor, reaching 5.4 ± 0.9 × 109 infectious viral particles (IVPs)/mL. Notably, in vitro neutralization assays using a Transwell co-culture system demonstrated a 4.3 log reduction in viral titers across genetically diverse PRRSV-2 strains, including VR2332, JXA1, JS07, and SH1705. Collectively, this study provides a robust platform for large-scale rAAV production and highlights the potential of SVR-based gene therapy to address the antigenic diversity of PRRSV-2.
Keywords: PRRSV; adeno-associated virus vector; broad-spectrum antiviral activity; insect cell bioreactor; soluble viral receptor fusions.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures

Similar articles
-
Fusion expression of the two soluble viral receptors of porcine reproductive and respiratory syndrome virus with a single adeno-associated virus vector.Res Vet Sci. 2021 Mar;135:78-84. doi: 10.1016/j.rvsc.2020.12.026. Epub 2021 Jan 2. Res Vet Sci. 2021. PMID: 33453552
-
Additive inhibition of porcine reproductive and respiratory syndrome virus infection with the soluble sialoadhesin and CD163 receptors.Virus Res. 2014 Jan 22;179:85-92. doi: 10.1016/j.virusres.2013.11.008. Epub 2013 Nov 15. Virus Res. 2014. PMID: 24246307
-
Recombinant adenovirus-delivered soluble CD163 and sialoadhesin receptors protected pigs from porcine reproductive and respiratory syndrome virus infection.Vet Microbiol. 2018 Jun;219:1-7. doi: 10.1016/j.vetmic.2018.04.006. Epub 2018 Apr 4. Vet Microbiol. 2018. PMID: 29778179
-
Porcine reproductive and respiratory syndrome virus vaccines: current status and strategies to a universal vaccine.Transbound Emerg Dis. 2014 Apr;61(2):109-20. doi: 10.1111/tbed.12016. Epub 2013 Jan 24. Transbound Emerg Dis. 2014. PMID: 23343057 Review.
-
Advancements in molecular design and bioprocessing of recombinant adeno-associated virus gene delivery vectors using the insect-cell baculovirus expression platform.Biotechnol J. 2021 Apr;16(4):e2000021. doi: 10.1002/biot.202000021. Epub 2021 Jan 25. Biotechnol J. 2021. PMID: 33277815 Review.
References
-
- Neumann E.J., Kliebenstein J.B., Johnson C.D., Mabry J.W., Bush E.J., Seitzinger A.H., Green A.L., Zimmerman J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005;227:385–392. doi: 10.2460/javma.2005.227.385. - DOI - PubMed
Grants and funding
- 2024rc21, 2024rc26/This work was supported by the High-level Talents Scientific Research Initiation Project of Jiangsu Vocational College of Agriculture and Forestry
- 2018YFC0840400-4/he National Key Research and Development Program of China
- PAPD/Priority Academic Program Development of Jiangsu Higher Education Institutions
LinkOut - more resources
Full Text Sources
Research Materials
